Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3713
(Return to index)
Principles of Physiological Psychology
By Wilhelm Wundt (1902)
Posted March 2000
[p. 104] CHAPTER IV
Morphological Development of the Central Organs
§1. General Survey
(a) -- Object of the Following Exposition
In the preceding chapter we have attempted to analyse, in their elementary phenomena, the vital processes conditioned upon the constitution of the nervous substance. Now in every organism which stands high enough in the scale of organic life to possess a nervous system at all, the elementary parts connect to form complicated structures, or organs; and the processes which we have been studying manifest themselves, accordingly, in co-ordinated activities of greater or less complexity. As a rule, it is far from easy to refer these complex phenomena to the relatively simple conditions laid down by a mechanics of nerve-substance that has been worked out from individual, isolated structures. We must he content to do this in the rough, and as a matter of general direction. At the same time, the more complicated and difficult the problem, the more strictly necessary is it, if our analysis of the complex physiological functions is not to go wrong from the very beginning, to keep constantly in view the general principles which our study of the simple nerve processes has brought to light. In considering, more especially, those developmental forms of the nervous system which are of chief importance for psychology, and which will therefore form the main subject of the following Chapters, -- the nerve-centres of the higher vertebrates and of man, -- we must remember that the elementary nerve-forces are still at work, though the manifold connexions of the elementary parts place their effects under conditions of almost inconceivable complexity. The properties of compound organs can be understood only in so far as we are able to refer them, at any rate as regards the general point of view from which they are appraised, to the properties of the elementary structures. The more firmly the physiology of the nervous system holds to this principle, which is surely beyond the need of argument, the sooner will it be competent to render service to psychology. On the other hand, neglect of the rule, combined with the adoption of a haphazard popular psychology, as manufactured on occasion by anatomists and physiologists for their own [p. 105] private use, has wrought havoc from the days of GALL and phrenology down to the present time. Great advances have been made in our knowledge of the morphology of the nervous system and of its complex physiological functions; GALL himself deserves credit for his investigations in anatomy. But they have failed to bear fruit for an understanding of the relations of the nervous system to the processes of the psychical life. Nay, more, under the conditions just mentioned, they have in this regard oftentimes done more to confuse than to further knowledge.
Now an inquiry concerning the bodily substrate of the mental life evidently presupposes, first of all and before it turns to the properly physiological aspect of its problem, an adequate knowledge of the morphology of the organs. This is not a merely logical requirement. Apart from the experiments on nervous elements, isolated so far as possible from their connexions, which fall within the sphere of the general mechanics of nerve-substance, both the physiological experiment and the pathological observation which serves to supplement it in various directions are conditioned upon this anatomical knowledge. When we remember the immense complexity of the structures involved, we must admit, at the same time, that as instruments of analysis they are comparatively crude: a limitation that should be steadily borne in mind in any estimate of the value of such experiments and observations. The first part of the following discussion will, then, be devoted to a general sketch of the morphology of the central nervous system. This will, of course, contain nothing that is new to the anatomist and physiologist, to whom the subject is familiar. On the contrary, it will often fail to make mention of special points that, for the time being, possess only an anatomical interest. The primary purpose of the exposition is to furnish a brief account of the structure and function of the nervous system that shall appeal especially to the psychologist and shall take account of the things that interest him. It has, however, over and above this, the secondary purpose of showing the anatomist and physiologist themselves how the familiar facts of structure and function appear when viewed, for the nonce, from the standpoint of psychology. It is true that this standpoint is not altogether neglected in the anatomical and physiological text-books. On the contrary, one cannot but admire the courage with which the anatomists and physiologists, when occasion arises, invade a difficult and (for so it really is to them) an unknown country. But the psychologist is, all the more for this very reason, in duty bound to discuss, from the point of view of a scientific psychology, the results obtained from the observation of microscopical structures, or of animals whose mind has in some way been impaired by extirpation of particular portions of the brain. He is bound to pass the facts themselves in critical review, and to estimate their psychological [p. 106] importance, quite apart from the more or less accidental and arbitrary reflections that are usually appended to the observational data. Now it would obviously be out of place, for the purposes of a survey specially intended, as this is, to subserve psychological requirements, to give any such detailed account of the topography of brain structure as is necessary for anatomy and pathology. We must rather lay the chief emphasis upon the morphological complexes, upon the organs as such; more particularly where structural connexions point us to co-ordination of functions. And this need seems to be most adequately met by a genetic consideration, which seeks to explain the complex conditions found in the fully developed organs from their origination in simpler forms, whether these stand lower in the scale of organic life or represent earlier stages of individual development. We shall, therefore, in this opening Chapter, say what is necessary for our present purpose of the general differentiation of the substrate of psychical functions in the animal kingdom. That done, however, we may thenceforth confine ourselves to a detailed consideration of the morphological development of the central organs in the vertebrates. Here, too, we shall make use of the lower forms of development chiefly to prepare the way for an understanding of the structural plan of the human brain.
(b) -- The Neural Tube and the Three Main Divisions of the Brain
The first step in the evolution of the central nervous system of vertebrates is taken, as we have seen, with that primitive differentiation of the germ where a dark streak of tissue marks the place of the myel, and therefore the longitudinal body-axis of the future organism (Fig. 8, p. 37). The next stage in the development of this primule of the nervous system occurs when the outer layer of the germ disc folds on either side of the axis of the primitive streak to form two ridge-like elevations, containing a groove between them. This, the primitive groove, is the primule of the future myel. The sides at first grow rapidly upwards, and then bend inwards, so that the groove closes to form a tube, the neural tube, within which the myel arises by proliferation of the original formative cells (Fig. 9, p. 37). The primary cavity of the myel persists throughout the vertebrate series as the longitudinal central canal or myelocele. This is lined with cinerea or grey matter, which is itself invested with an envelope of alba or white matter; and from this, again, the roots of the myelic (spinal) nerves issue in a fan-shaped radiation.
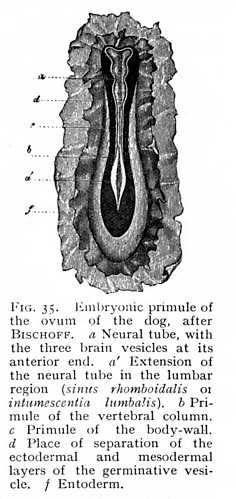
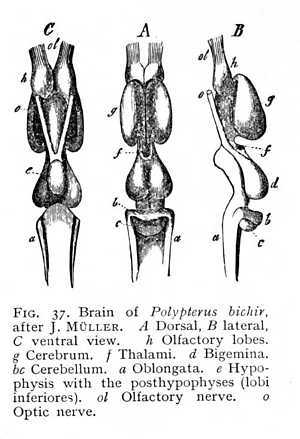
The primule of the brain is formed from the anterior end of the neural tube. The rapid growth of this region leads to the formation of a bladder-like expansion, the primitive brain vesicle, which soon divides into three compartments, the fore, mid and hind brain-vesicles (Fig. 35). Both the genetic and the later functional relations of the primary vesicle suggest that this tripartite division, like the development of the brain at large, is intimately connected with the development of the three anterior sense-organs. The nervous primule of the olfactory organ grows out directly from the anterior end of the fore brain; that of the auditory organs from the lateral walls of the hind brain; while, despite the fact that the eyes would seem to represent product of the growth of the fore brain, we must, in the light of indisputable physiological facts, look to the mid brain as the ultimate source of origin of the optic nerves.
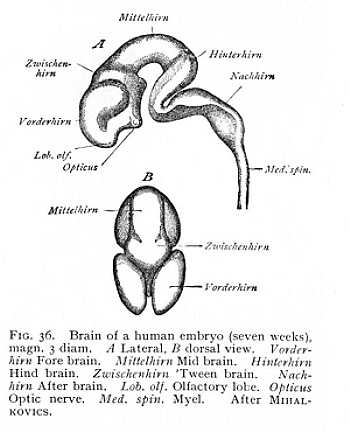
Of the three primary brain-divisions: the first and third, the fore and hind brains, undergo the greatest changes. The anterior
extremity of both soon begins to outstrip the rest in growth, so that both alike divide into a principal and a secondary vesicle. The original fore brain now consists of fore brain and 'tween brain, the original hind brain of hind brain and after brain (Fig. 36). Of the five brain divisions which have thus arisen from the three primary vesicles, the fore brain or prosencephalon corresponds to the future cerebral hemispheres; the 'tween brain or diencephalon becomes the thalami; the undivided mid brain or mesencephalon developes into the quadrigemina of man and the animals, the bigemina or optic lobes of the lower vertebrates; the hind brain or epencephalon becomes the cerebellum; and the after brain, or metencephalon, the oblongata. The diencephalon is to be considered the anterior, and the metencephalon the posterior stem-vesicle, from which the prosencephalon and epencephaon have grown out respectively as secondary vesicles. The structures developed from the three stem-vesicles (metencephalon, mesencephalon and diencephalon), -- i.e., the oblongata, the quadrigemina and the thalami, -- and the fibre-systems that ascend among them from the myel, are grouped together in the nomenclature of the developed brain as the caudex or brain-stem. The structures of the first and fourth vesicles, the cerebral hemispheres and cerebellum, are named, in contradistinction to the brain-stem, the pallium or brain-mantle, since in the more highly organised brains they envelope the brain-stem as a mantle-like covering.[p. 108]
The three brain vesicles, then, represent expansions of the anterior end of the neural tube. With the tube itself, they form a
closed system, whose parts intercommunicate by way of the continuous central cavity, But the development of the two secondary vesicles from the first and third primary vesicles brings other changes with it. The roof of the fore and hind brains divides up longitudinally; so that two slit-like apertures appear lying exactly in the median line of the body, whereby the cavities of the fore and hind stem-vesicles are exposed. The anterior roof-slit divides the prosencephalon into its two hemispheres, and leaves the diencephalon open above. The mesencephalon, which does not share the advance in organisation characteristic of the rest of the brain, merely divides by a longitudinal furrow into two halves. The posterior roof-slit appears at the place where the neural tube passes over into the brain. The cerebellum, which grows out directly forwards from this point, is at first separated into two entirely distinct halves, but afterwards grows together again in the median line. The two roof-openings serve to admit blood-vessels into the brain-cavities. These ensure the food supply requisite for further growth and for the simultaneous thickening of the brain-walls by deposition of nerve-substance from within.
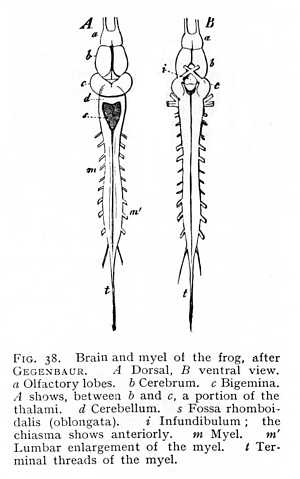
The level of development now attained is practically the level of permanent organisation in the lowest vertebrates, the fishes and amphibia (Figg. 37, 38). The original prosencephalic vesicle is here divided, in most cases, into two almost entirely separate halves, the cerebral hemispheres; the only remaining connexion occurs over a small area of the floor of the vesicle. The anterior stem-vesicle (diencephalon) is split into two lateral halves, the thalami, which retain connexion at their base. The cerebellum forms for the most part a narrow unpaired lamella, from which [p. 109] all trace of division has disappeared. In the metencephalon or oblongata., the posterior roof-slit has formed a rhomboidal depression, on the floor of which the principal mass of the organ shows in undivided form.
(c) -- The Brain Ventricles and the Differentiation of the Parts of the Brain
The division of the brain into five vesicles brings with it a further change: a modification in the form of the central brain cavities, whose origin as simple expansions of the myelocele we have already noticed. The brain-cavity divides, in accordance with the
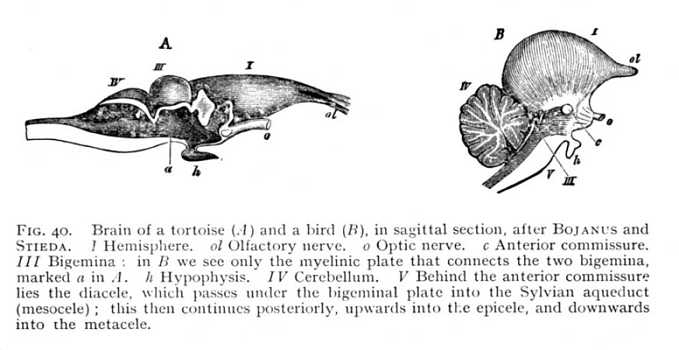
separation of the brain-vesicles, at first into three and then into five pockets. The division of the hemispheres subdivides the first of these again into two symmetrical halves, the paraceles or lateral brain-ventricles. If, now, we set out from these on a journey through the brain-cavities, we shall traverse them in the following order (Fig. 39). The two paraceles (h), which as a rule are entirely separated from each other, open into the cavity of their stem-vesicle, -- a cleft-like space, bounded laterally by the thalami, and left roofless by the anterior roof-slit (z), -- the diacele or third ventricle. This leads directly into the cavity of the mesencephalon (m). In mammals the mesocele is extraordinarily [p. 110] reduced, so that it appears only in the form of a narrow canal, the aqueduct of Sylvius, running below the quadrigemina, and connecting the diacele with the cavity of the metencephalon. In the birds, the mesocele is more extensive, sending lateral offshoots into the bigemina (mesencephalon); and in the lower vertebrates
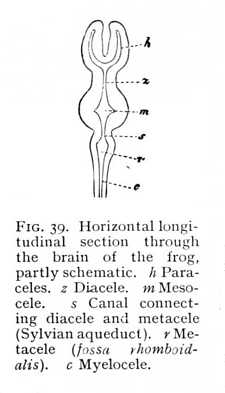
the bigemina contain quite large cavities, which communicate with the central cavity. The two derivatives from the third primary vesicle, epencephalon and metencephalon, have originally each its own special cavity. But the cerebellum (epencephalon) is now a rounded vesicle, arching backward over the metencephalon front the point at which this borders upon the mesencephalon. The mesocele accordingly divides at its posterior end into two branches, the one of which turns upwards, and leads into the epicele (cavity of the cerebellum), while the other pursues a straight course into the cavity of the metencephalon or oblongata (Fig. 40). This latter cavity, the metacele (fourth ventricle [1]) is termed from its rhomboidal form the fossa rhomboidalis (r in Fig. 39). The metacele is, therefore, not strictly a cavity, but a furrow; the posterior roof-slit has completely exposed it. It closes posteriorly to pass over into the myelocele. In mammals the epicele disappears entirely by the filling up of the epencephalic vesicle with alba. At this stage, then, the paraceles,[p. 111] diacele, Sylvian aqueduct (mesocele) and metacele represent the whole system of brain-cavities. In the lower vertebrates, we have, further, the cavities of the thalami as extensions of the diacele; the cavities of the bigemina as branches of the aqueduct; and the epicele as a tributary of the metacele. In the lower orders of vertebrates, primary and secondary cavities alike are, as a general rule, more extensive in proportion to the mass of the brain, -- i.e. approximate more nearly to an embryonic state. However, the different classes evince wide differences in this regard as between the various subdivisions of the brain. In the fishes, the cerebral hemispheres and cerebellum are filled with alba and become solid structures. Their growth is soon arrested, and they consequently attain to no considerable size. In the amphibia, the paraceles persist, but the cerebellum is
usually solid. When we come to the reptiles and birds, we find a cerebellum with a spacious epicele; but in all the mammals this has again

disappeared. In the mammals, too, the lateral cavities of the mesencephalon (quadrigemina or bigemina) are lost; in all the lower vertebrates, from the fishes to the birds, they not only persist but develop prominences of cinerea upon their floor (Fig. 41). Similar growths, the striata, appear in the paraceles of the avian and mammalian brain.
Both in the myel and in the brain (encephalon), the nervous mass is formed by proliferation of the cells constituting the walls of the original cavities. Many of these cells evince the character of the formative cells of tissue, and so mediate the secretion of the amorphous intercellular substance or neuroglia. Others become nerve-cells, and send out runners. In the myel, the greater part of the fibres radiate out towards the periphery, so that the cinerea is collected about the myelocele and surrounded by an envelope of alba. In the encephalon this distribution of alba and cinerea persists, practically unchanged, in the structures developed from the three stem-vesicles. In the developments from the secondary vesicles, on the other hand, the nerve-cells retain their position in the walls of the cavities, and the fibres connected with them trend towards the interior. Hence in the caudex -- the oblongata, the quadrigemina and the thalami -- we have a layer of cinerea lining the continuations of the myelocele and surrounded by an envelope [p. 112] of alba; in the pallium, a mass of alba invested with a covering of cinerea. The Linerea thus shows two distinct formations. The one, the entocinerea (tubular grey matter) belongs to myel and caudex, the other, the ectocinerea (cortical grey matter), to the pallium. The entocinerea of the encephalon undergoes still further transformation. Even in the highest region of the myel, various bundles of nerve-fibres from the myelic columns have shifted from their former position at the periphery of the cenerea, so that this is broken up by masses of alba. In the oblongata, this process has gone so far that only a comparatively small part of the cinerea holds its original place as floor of the fossa rhomboidalis, by far the greater portion being separated by the intercurrence of myelinic fibres into distinct masses. These
collections of entocinerea, invested by alba, are termed nidi (nuclei). We see, then, that the entocinerea of the myel undergoes an essential modification as it passes into the encephalon; it is broken up by the interposition of masses of alba, and so gives rise to a third formation of grey matter, the nidal cinerea (nuclear grey, ganglionic grey). This nidal cinerea lies midway between the entocinerea and the ectocinerea. As we travel from the central cavity towards the periphery, we come first of all upon entocinerea, then upon the nidal cinerea, then again upon alba, and finally upon the ectocinerea.
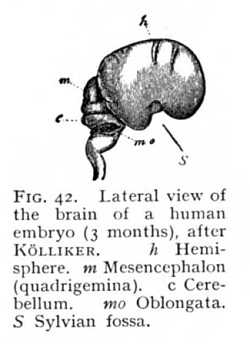
The series of changes that we have been describing hitherto is accompanied, in all the vertebrates, by changes in the relative position of the primitive brain-divisions, as a result of which the whole brain is bent over ventralward. The various parts of the caudex are thus brought out of the straight line and set at a certain angle to one another. The bend or flexure, which in the lowest classes is but slight, approximates more and more closely to a right angle the higher we ascend in the vertebrate series (Fig. 36, p. 180). And the form of the brain is further modified by the disproportionate growth of certain divisions, especially the prosencephalon and epencephalon, which extend over and conceal the rest. Three flexures of the central nervous system can thus be distinguished. The first appears at the junction of the myel and oblongata; the second takes place in the cerebellum; and the third at the level of the mesencephalon (Fig. 42). The extent of these flexures is principally conditioned by the growth of the prosencephalon; so that degree of curvature runs practically parallel with development of the hemispheres.[2][p. 113] In the early stages of the development of the vertebrate brain, the prosencephalon extends anteriorly beyond the remaining brain-divisions, without covering them. But the more its growth outstrips the growth of the rest of the brain, the more opposition does the rigid attachment of the embryo to the germinative vesicle offer to its forward expansion. It must, therefore, grow out backwards, arching first over the diencephalon, then over the mesencephalon, and finally over the cerebellum itself. At the same time, it follows the curve of the mesencephalic flexure; its most posterior portion -- that which covers mesencephalon and epencephalon -- bends upon itself at an angle. The more vigorous the growth of the hemisphere, the farther does the bent portion of it extend back again towards its point of departure, or, in other words, the more nearly does the curve described
about the diencephalon approach to a perfect circle. In this way a depression (the Sylvian fossa; s Fig. 42) is formed at the place where the hemisphere rests upon its stem-vesicle (diencephalon). In the most highly developed mammalian brains, the concavity of the curve of growth is almost entirely obliterated, and the walls of the fossa draw together, leaving between them a deep and narrow fissure.
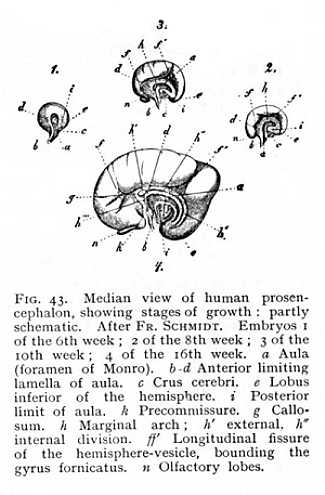
The growth of the prosencephalon over the caudex is necessarily followed by a modification of the form of the paraceles. These are originally spherical cavities, lying within the hemispheric vesicles. As the prosencephalon grows, they extend out at first posteriorly, and then, when the curve of the hemispheric arch begins to close in upon itself, inferiorly and anteriorly as well. The central portion of the resulting cavity is termed the cella; the extensions are the postcornu, precornu and medicornu of the paracele. While this transformation is in progress, the outer walls of the prosocele are growing more rapidly than the inner or median wall which surrounds the caudex. In this there is a narrow, [p. 114] originally vertical slit, the aula or foramen of Monro (a Fig. 23) whereby the paraceles communicate with the diacele (third ventricle). Anteriorly to the aula, the hemispheres are held together by a lamella of alba (b-d). As the prosencephalon arches over the rest of the brain, the aula and its anterior limiting lamella naturally arch with it. They have, therefore, in the developed brain, the appearance of a vault laid over the diencephalon. The posterior part of the aula soon closes, and only the extreme anterior portion of the original cleft remains open; this serves as a channel for vascular processes passing from the diacele into the paraceles. The lower end of the lamella of white matter, which forms the anterior boundary of the aula, becomes the precommissure (k); the remaining portion follows the curve of the hemispheric arch and is the primule of the fornix. Directly above this the hemispheres are united by a strong transverse band of alba, the callosum or great commissure (g). The portion of the median wall of the hemispheres lying above the callosum forms yet another arch, running concentrically with the fornix, and separated from the surrounding parts by a special furrow, f f'. This is the marginal arch, h. Its anterior division becomes the gyrus fornicatus; the posterior passes into a structure, continuous with the gyrus fornicatus and extending from the median wall outwards into the paracele, termed the hippocampus (cornu Ammonis). A more detailed description of these parts, which attain to development only in the mammalian brain, will be given later.
§2. The Myel in the Higher Vertebrates
The neural tube from which the myel developes is originally a hollow tube, filled with liquid, and lined along its interior wall with
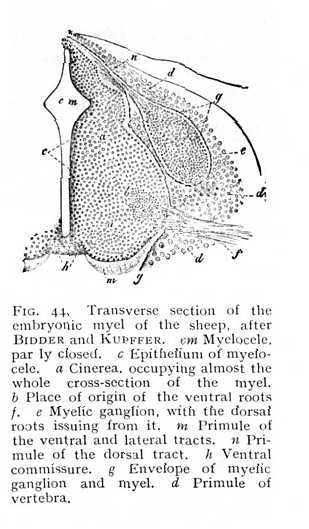
formative cells. The cells increase and multiply: some taking on the character of connective tissue-cells, and furnishing an amorphous intercellular substance, while others become nerve-cells. The processes of these last either pass directly into the fibres of peripheral nerves, or divide and subdivide to form a terminal reticulum. The main trend of all fibres is to the periphery of the neural tube, so that the cellular structures are shifted towards the centre of the myelic cavity (Fig. 44, and Fig. 9, p. 37 above). The nerve-cells, and the nerves issuing from them, are arranged, from the first, in accordance with the bilateral symmetry of the primule of the vertebrate body, in symmetrical (right and left) groups. Moreover, the connexion of the nerves with two different parts of the germ-primule carries with it the further separation of each group into two subdivisions. All nerves and fibres that enter into connexion with the corneal layer, the primule of the sense-organs and the sensitive investment of the body, arrange themselves in a dorsal group, in the near neighbourhood of the germinal structures dependent upon them.[p. 115] All those nervous elements, on the other hand, that have relations to the striated muscles collect in a ventral group, corresponding to the animal muscle-plate. It results from this that the cinerea formed by the association of cells appears to right and left as a dorsal and a ventral column, surrounded by an envelope of myelinated fibres or alba. The columns are termed, from their appearance in transverse section, the dorsal and ventral cornua (horns) of the myel: a special branch of the latter is known as the lateral cornu. The dorsal and ventral cornua of each side are united at the centre. The nerve-roots issuing from the cornua are arranged, in the same way, in two series: the dorsal or sensory, and the ventral or motor (Fig. 44, e, f; Fig. 45, HW and VW).
Under these conditions of growth, the myelic cavity at first assumes the form of a rhombus (Fig. 44, cm), drawn out into a ventral and a dorsal cleft. The cavity of the dorsal cleft soon becomes almost entirely filled; that of the ventral is more plainly
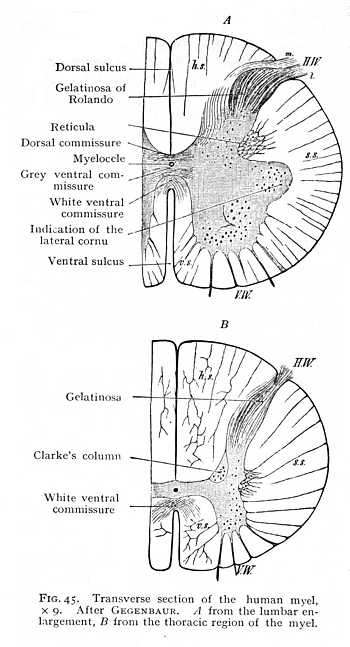
marked, but is closed by nerve-fibres running from side to side of the myel, and constituting the white ventral commissure. The commissure, which originally crosses near the periphery (Fig. 44 h), gradually reaches a deeper level (Fig. 45). Behind it, a remnant of the myelic cavity persists as an extremely narrow canal, the myelocele or central canal of the myel, around which the two collections of cinerea enter into cross-connexion (Fig. 45 A). The dorsal and ventral sulk (fissure median post. and ant.) divide the myel into two symmetrical halves. Each of these is again subdivided by the outgoing nerve-roots into three columns (Fig. 45 B). The column of alba lying between the dorsal sulcus and the cell-column of the dorsal roots is termed the dorsal column (hs); that lying between the ventral sulcus and the cell-column of the ventral roots the ventral column (vs); and a third column, ascending between the cell-columns of the dorsal and ventral roots, the lateral column (ss). The greater part of the nerve-fibres of these white columns run their course vertically, in the direction of the longitudinal axis of the myel. An ex-[p. 116]ception to the rule is furnished by the area at the central end of the ventral sulcus, occupied, as we saw just now, by the ventral commissure; here the decussating fibres follow a horizontal or oblique course; and the same directions are, naturally, taken for a short distance by the fibres which constitute the direct continuations centralward of the incoming nerve-roots. The grey cornua are of different shapes: the ventral, particularly in the lumbar region of the myel, are broader and shorter, the dorsal longer and narrower. The former contain a quantity of large multipolar ganglion-cells; the latter consist exclusively of smaller cells. A good portion of the dorsal cornua consists, further, of the nervous ground reticulum and its interpenetrating fibrils. This gives them a peculiarly bright appearance, more especially as they approach the periphery of the myel: the region is known as the gelatinosa of Rolando. Passing centralward from this formation we find on either side of the central canal a compact column of alba and cinerea, Clarke's column (Fig. 45 B),[p. 117] containing a well-marked group of roundish ganglion-cells, and extending from the end of the cervical region into the lumbar enlargement. The immediate points of origin of the dorsal roots, within the myel, seem to be less richly supplied with nerve-cells than those of the ventral; but the difference is compensated later on. A cluster of large bipolar ganglion-cells is intercalated in the course of the nerve-fibres after they have left the myel, and forms with them the spiral ganglion of the dorsal roots (e Fig. 44). The dorsal columns are mot connected, as the ventral are, by white myelinated fibres, but by a grey commissure, composed of fine fibres running transversely in the mass of cinerea behind the myelocele (dorsal comm., Fig. 45 A). Similar grey fibres surround the whole of the central canal; its interior is lined by a single layer of cylindrical epithelium, derived from a small remnant of the formative cells that originally invested the cavity of the neural tube (Fig. 9, p. 37 above).
So long as the development of the central organs is confined to the formation of the myel, we find, of course, a certain uniformity dominating the entire bodily organisation. The myel, over its whole length, simply repeats the same arrangement of elementary parts and the same mode of origin of nerve-fibres; and the sensory surfaces and motor apparatus that depend upon it must accordingly show a like uniformity of distribution and structure. Hence we find, as a matter of fact, that so long as the central nervous system of the embryo consists merely of the neural tube, no one of the higher sense-organs attains to development. The primules of the sensory investment of the body and of the locomotor apparatus are distributed uniformly about the central axis. A single exception occurs at the place where the nerve-mass takes on a stronger growth, for the supply of the posterior extremities; this is indicated, at an early stage, by an extension of the primitive groove, the sinus rhomboidalis or future lumbar enlargement. A similar, but less strongly marked thickening of the neural tube occurs, later on, at the point of departure of the nerves supplying the anterior extremities, -- the cervical enlargement.[3] This uniformity of organisation is permanently retained in the lowest vertebrate, Amphioxus lanceolatus, in which the development of a central nervous system halts at the formation of the neural tube. The organ of vision, in this brainless vertebrate, consists of simple refracting cells, surrounded by pigment [4]; the organ of smell, of an unpaired cup-shaped depression at the anterior end of the body; the presence of an auditory organ has not been demonstrated. We see, then, that the arrest of development in this case affects just those [p. 118] organs which appear to exercise a determining influence upon the formation of the higher central parts, the vesicular differentiations of the myel.
§3. The Oblongata
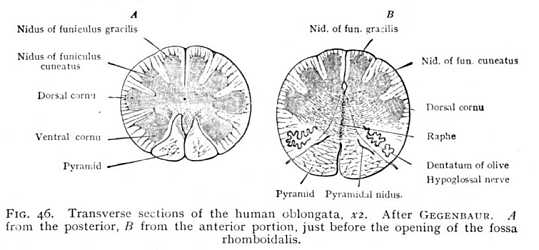
In the lower vertebrates, the bundles of nerve-fibres take a course that is, to all appearance, but little different from their course in the myel. The only changes are that the dorsal columns split apart, disclosing the fossa rhomboidalis (Fig. 37, 38, p. 109);
and that the grey cornua, as may be seen in section, have been divided off from the central cinerea and intercalated in the course of the ventral and dorsal columns. The three columns of the myel, ventral, lateral and dorsal, can still be distinguished,
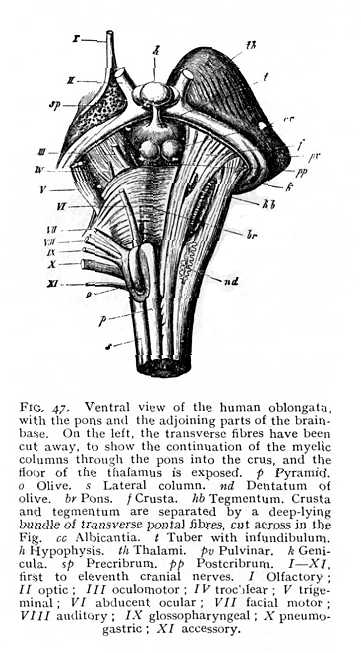
but receive special names. Their fibres pursue a more tortuous course, and ganglionic nidi appear among the fibre-bundles. They thus differ very considerably from the correspondingly situated myelic columns; and, indeed, for the most part do not represent direct continuations of the myelic structure. The ventral columns are termed pyramids; their fibres decussate, in the posterior part of their course, so that the ventral sulcus is entirely obliterated (Fig. 46 A; Fig. 47 p). This decussation seems to be a repetition, on a larger scale, of the decussation of the ventral columns of the myel in the ventral commissure. At their upper end, where they enclose a ribbon-like stria of cinerea (pyramidal nidus, inner accessory nucleus of olive; pyram. nidus, Fig. 46 B), the pyramids are bounded on either side by the olives (Fig. 46 B; Fig. 47 o). These are well-marked prominences, which contain in their interior a large ganglionic nidus, dentate in section and therefore termed the dentatum (nd). The vertically ascending fibre-bundles which enclose the dentate nidus are known as the capsular columns (finiculi siliquae). The lateral columns (s Figg. 47, 48) grow smaller and smaller [p. 119] from the lower end of the oblongata upwards, until at about the point where the opening of the fossa rhomboidalis appears they disappear entirely in the interior. The dorsal columns, on the other hand, increase in external diameter. In the lower portion of the oblongata, they are divided by a shallow sulcus into inner and outer columns, the slender and the cuneate funiculi (fg and fc Fig. 48), which at the lower end of the fossa rhomboidalis carry knob-like prominences, produced by the grey nidi of the interior (nid. grac. and nid. cun., Fig. 46). Farther upwards, the two funiculi appear to continue their course in the columns which bound the fossa rhomboidalis to right and left. These have been termed the restes (restiform bodies: pi Fig. 48). They are the largest columns of the oblongata, and, like the funiculi just mentioned, contain grey nidi. They are characterised by the intricate, trellis-like interlacing of their constituent fibres. Anteriorly, the restes pass entirely over into the alba of the cerebellum forming the inferior cerebellar peduncles. Between them, on the floor of the fossa rhomboidalis and directly covered by entocinereal matter, are two further tracts, which appear to represent the continuations of the parts of the myel lying ventrally of the myelocele, i.e. the ventral cornua and the deeper-lying portions of the ventral columns. These structures, which extend over the whole floor of the fossa rhomboidalis, and are principally [p. 120] composed of cinerea, are known, from the arching convexity of their form, as the cylindrical columns (eminentiae teretes, et). Their cinerea is in connexion with most of the grey
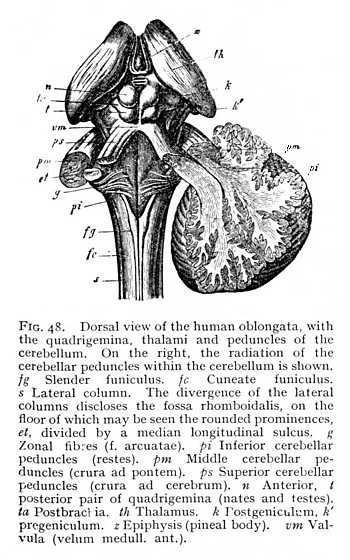
nidi of the oblongata, though some of these are forced out from the median line, and thus isolated, as a result of the splitting up of the oblongata by tracts of alba. A further and final consequence of the changes of structural conditions which we have been describing is the formation of an entirely new system of fibre-groups surrounding the oblongata in a transverse direction. Some of them enter into the ventral sulcus and the sulcus separating the pyramids from the olives; others strike across the fossa rhomboidalis. The system is known as the zonal fibre-system (stratum zonale, fibrae arcuatae, g).
The re-arrangement of parts in the oblongata leads to a redistribution of the points of origin of the peripheral nerves. The simple rule of the myel is no longer obeyed, the nerve-roots are more or less displaced from their old positions. It is true that they still form, roughly, two longitudinal series, a dorsal and a ventral. But root-fibres that are exclusively motor issue only from the ventrolateral sulcus (twelfth cranial nerve, hypoglossal or lingual motor). The dorsolateral sulcus (or, at any rate, its immediate neighbourhood) gives rise, on the other hand, both to sensory and to motor fibres. Here begin all the other cranial nerves, with the exception of the first and second, olfactory and optic, and the third and fourth, the two anterior oculomotor nerves, whose place of origin lies further forward (cf. Figg. 47, 52).[5][p. 121]
§4. The Cerebellum
At the anterior end of the oblongata, the structural relations of the parts are further, and very essentially, modified, owing to the outgrowth of the cerebellum from the primule of the third brain-vesicle (epencephalon). At the lowest stage of its development, the cerebellum varies but little, in outward form, from its original primule (Figg. 37, 38, p. 109); it consists of a transverse stria,
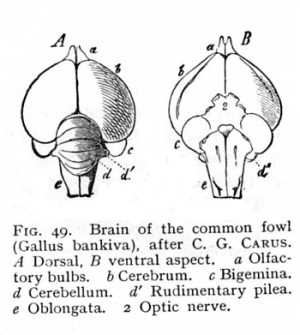
bridging over the anterior end of the fossa rhomboidalis and receiving into its substance the restis of either side. Anteriorly, it is continued by a myelinated plate to the mesencephalon (Fig. 40, p. 110); laterally, it gives out tracts of transverse fibres, which run towards the lower surface of the oblongata, and decussate with each other and with the vertically ascending fibre-tracts of the pyramidal and olivary columns. These connexions remain the same later on, when the cerebellum has attained its further development. The bundles that enter it from the restes are the inferior cerebellar peduncles (crura ad medullam oblongatam, pi Fig. 18); the myelinated fibres issuing from it towards the mesencephalon are the superior cerebellar peduncles (crura ad cerebrum, ps). These are united by a thin myelinated plate, the valvula (velum medullare superius, vm), which forms the roof of the fossa rhomboidalis, and effects a direct connexion of the cerebellum with the adjoining anterior brain-division, the mesencephalon or quadrigemina. Finally, the white tracts proceeding from either side of the cerebellum form the middle cerebellar peduncles (crura ad pontem, pm). The structure that arises at the brain-basis by the junction of the middle peduncles and their decussations with the longitudinal myelinated [p. 122] fibres ascending from the oblongata is known as the pons (pons Varolii, br Fig. 47). It constitutes on the one hand a connecting link in the longitudinal direction between metencephalon and mesencephalon, and on the other a connecting link in the horizontal direction between the two lateral halves (pilea) of the cerebellum. The superior and inferior cerebella-peduncles are clearly visible at the very earliest stage of cerebellar development. The middle peduncles, on the contrary, do not attain to a growth that enables us to distinguish the pons as a special structure until the development of the cerebellum, and especially of the pilea, has advanced a considerable distance. Even in the birds, their place is supplied by little more than the longitudinal continuations of the ventral and lateral columns of the oblongata (Fig. 49 B). From the points at which the peduncles enter the cerebellar
mass, above, below and sideward, myelinated fibres radiate out towards the periphery of the organ.
The morphological development of the cerebellum, the posterior section of the pallium, is completed at a comparatively early period. In all vertebrates, it is covered by ectocinerea, clearly differentiated from the radiation of myelinated fibres that occupies the interior; and even in the lowest vertebrates, the fishes, the cerebellar cortex divides into a number of layers, characterised by differences of coloration. In the cerebellum of the amphibia, we find groups of nerve-cells intercalated in the course of the myelinated fibres, -- the first traces of ganglionic nidi. In the birds, these increase in number, while at the same time the layer-formation of the cortex becomes plainer, and an increase in the mass of the cortical elements is rendered possible by superficial folding (Figg. 40, 49).
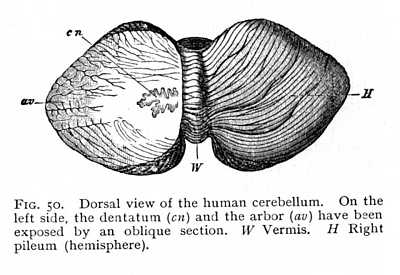
The cerebellum undergoes its final stage of structural development in the mammals. Here we find, first, an unpaired median portion, the surface of which is crossed by transverse folds, and which has received the descriptive name of vermis; and, secondly, more strongly developed symmetrical lateral portions, the pilea. In the lowest mammals, it is true, the growth of the vermis exceeds that of the pilea; but in the higher forms, the pilea grow out about it in all directions (Fig. 50). The development of the pilea is accompanied by a more vigorous growth of the middle peduncles, which in the lower vertebrates are merely indicated by slender transverse fibre-tracts to the oblongata. The transverse folds of the ectocinerea increase in number, and show in cross-section the well known dendritic appearance of the arbor (arbor vitae, av Fig. 50). At the same time, larger ganglionic nidi appear in the radiation of myelinated fibres within the cerebellar mass. Thus each pileum contains a dentate nucleus, similar to that of the olives (nucleus dentatus cerebelli, cn). Other cinereal nidi, analogous in function, are scattered among the alba of the pons; their cells are intercalated between the decussations of the various fibre-bundles.[p. 123]
§5. The Mesencephalon
The mesencephalon -- the division of the caudex which corresponds to the bigemina (lobi optici) of the lower vertebrate; and the quadrigemina of mammals (t, n Fig. 48; d Fig. 37) -- contains two formations of grey matter, entocinerea and nidal cinerea; only the secondary vesicles develope the ectocinereal pallium. The entocinerea surrounds the Sylvian aqueduct in a layer of
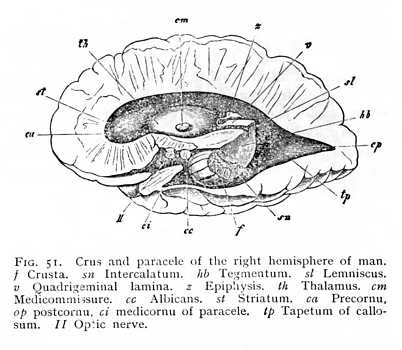
moderate thickness; the most anteriorly situated nerve-nidi (those of the oculomotor and trochlear nerves and of the upper root of the trigeminus) are in connexion with it. Ganglionic nidi are found scattered through the bigemina or quadrigemina, and intercalated in the course of the myelinated fibre-tracts that pass below the Sylvian aqueduct. These are paired tracts of alba, united in the median line, and forming in the first place continuations of the ventral and lateral columns of the oblongata. Anteriorly, they are strengthened by the addition of longitudinal fibre-tracts, proceeding from the quadrigemina and thalami. They are termed, over their whole extent from oblongata to cerebral hemispheres, the brain crura (crura cerebri). The mesencephalic portion of the crura in the mammalian brain contains two well marked ganglionic nidi, the one of which, characterised by its dark coloration, is known as the intercalatum (substantia nigra of Soemmering: sn Fig. 51). It divides the crus of either side into a posterior and exterior portion, the crusta (basis pedunculi, pes cruris cerebri; f Figg. 47, 51), and an anterior and median portion, the tegmentum (tegmentum pedunculi; hb Figg. 47, 51). The most anterior extremity of the median portion, a band of alba which curves anteriorly into a fillet, and directly underlies the quadrigemina, is termed the lemniscus (ribbon; or laqueus, fillet; sl Fig. 51). A second tegmental nidus is named from its colour the [p. 124] rubrum (nucleus tegmenti; hb Fig. 56). Dorsal of the crura lie the quadrigemina (v Fig. 51). Posteriorly, they are connected with the superior cerebellar peduncles; anteriorly and laterally, they give out myelinated fibres, which in part mingle with the fibres of the tegmentum, in part pass into the thalami, and in part form the fibres of origin of the optic nerves. The connexion with thalami and optic nerves is mediated, in the mammalian brain, by the quadrigeminal brachia (ta Fig. 48). The pregemina are joined by the prebrachia to the thalami, and the postgemina by the postbrachia to the postgeniculum. In the space between the pregemina and the posterior extremity of the thalami lies the epiphysis (conarium, pineal body; z Figg. 48, 51), which Descartes, in the old days, looked upon as the 'seat of the mind.' It is a highly vascular structure, which genetically represents, in all probability, a rudimentary sense-organ: it is supposed to be the central remnant of a median eye, functional in the primitive vertebrates. The mammalian quadrigemina are, as we have already seen (p. 111), completely solid. They are connected by a lamina of alba, which posteriorly forms the direct continuation of the valvula, and anteriorly passes into the postcommissure (cp Fig. 53) running along the boundary-line of quadrigemina and thalami.
§6. The Diencephalon
The diencephalon, or region of the thalami, is smaller than the mesencephalon in all the lower vertebrates (f Fig. 37, p. 109); in the mammalian brain their relation is reversed (th Figg. 47, 48, 51). In the fishes, however, we have an indication of the change: a paired continuation of the diencephalon extends posteriorly to the base of the brain, and there appears in the form of two hemispherical prominences, lying ventrally and somewhat anteriorly of the bigemina (lobi optici). These are the inferior lobes of the fish-brain (li Fig. 41, p. 111). They enclose a cavity, which stands in connexion with the diacele, the cleft-like aperture
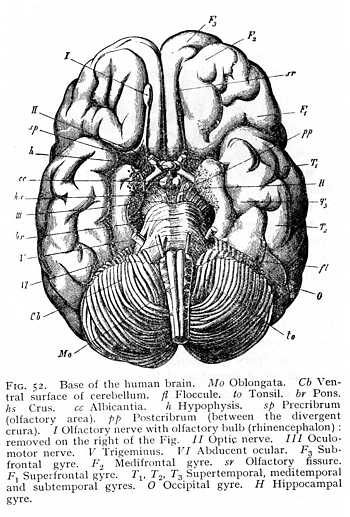
resulting from the anterior roof-slit and dividing the diencephalon into the two thalami. At the place where the inferior lobes meet in the median line, they are continued into an unpaired structure, the hypophysis (h Fig. 41), whose dorsal portion is an evagination of the diencephalon, while its ventral half is a remnant of embryonic tissue that originally belonged to the anterior extremity of the gullet, and remained in conjunction with the diencephalon when the base of the cranium developed. The hypophysis persists in the higher vertebrates, after the inferior lobes have entirely disappeared in consequence of the more vigorous development of the crura (h Fig. 52). The only point at which the ganglionic substance of the diencephalon appears, in these [p. 125] forms, is between the divergent crura, where we find a grey prominence, the tuber cinereum. This is continued anteriorly in the direction of the hypophysis, in a funnel-shaped prolongation, the infundibulum (i Fig. 38; t Fig. 47). The infundibulum contains a small cavity, communicating dorsally with the diacele. The entrance of small blood-vessels into the brain-mass gives the cinerea between the crura a perforated, sieve-like appearance; the region is termed the postcribrum (lamina perforata posterior; pp Figg. 47, 52). In the mammalian brain, two myelinated prominences, the albicantia (corpora candicantia or mammillaria; cc), issue ventrally from the floor of the diencephalon. Situated immediately in front of the anterior line of the pons, they bound the tuber posteriorly, as infundibulum and hypophysis bound it anteriorly. Their genetic signification is still uncertain.
The diencephalon, like the mesencephalon, contains two formations of grey matter, entocinerea and nidal cinerea. In the first place, the interior of the diacele is lined with a cinereal layer, which at the same time invests a thin band of alba, joining the two thalami, and termed the medicommissure (cm Fig. 51). The entocinerea of the diacele extends ventrally to the brain-[p. 126]base, where it is directly continuous with the tuber and infundibulum. Secondly, however, a number of ganglionic nidi, separated by masses of alba, are scattered throughout the interior of the thalami (th Fig. 56). Similar nidi may be found in two smaller rounded prominences, which in mammals form the posterior boundary of the thalami and externally are in connexion with them, the pregeniculum and postgeniculum (k k' Fig. 48, p. 120). The fibres of origin of the optic nerve interlace with the fibres of both genicula, and the postgeniculum alsohreceives [sic] the quadrigeminal postbrachium. The anterior and lateral portions of the thalami show a gentle roof-slope; posteriorly, the dorsal surface is separated from the ventral by a marginal swelling, the pulvinar (pv Fig. 47, p. 119)
§7. The Prosencephalon
(a) -- The Brain Cavities and the Surrounding Parts
In the earlier stages of its development, the prosencephalon is a vesicular structure, overlying the diencephalon. Originally simple, it is later divided by the anterior roof-slit into symmetrical halves, entirely separate save for the continuity of their floor. At the place where the roof-slit of the diencephalon is continued into the intercerebral fissure, the diencephalic cavity was
primitively in open communication with the two paraceles. In all the vertebrates (except the fishes, whose hemispheres are solid structures: p. 111), the vascular trunk that penetrates the cavity of the diencephalon sends out a large number of branches from it into the hemicerebral vesicles. When the diencephalon becomes so far solid, by growth of the constituent nerve-mass, that only the diacele (third ventricle) is left, the earlier doors of communication are almost entirely closed; only two very narrow apertures are left, at the anterior end of the diacele, which permit of the entrance of blood-vessels into the hemicerebral cavities. These are the portae (foramina of Monro: mo Fig. 53), the remnants of the original aula (Fig. 43, p. 113). They are separated anteriorly by a septum of alba, which represents the posterior line of junction of the two prosencephalic vesicles. The floor of the septum is usually formed of large bundles of myelinated fibres, transverse in direction, termed the precommissure (ca). In the reptilian, and still more in the avian and mammalian brains, the hemispheres come to such a growth as to arch more or less completely over the diencephalon. As a result of this, the paraceles run out posteriorly, and the thalami, instead of lying behind the hemispheres, as they did at first, form prominences that project with the greater part of their surface into the paraceles, and show only their internal faces to the diacele.[p. 127]
The grey matter of the prosencephalon occurs in all three possible formations. As entocinerea, it covers the walls of the diacele, and therefore, more especially, the inner faces of the thalami and the cavity of the infundibulum, as well as the whole of the infundibular region; as ganglionic cinerea, it forms considerable masses, intercalated in the course of the continuations of the crura below the thalami; and as ectocinerea, it invests the hemicerebral pallium at large. The position of these collections of grey matter, and their relation to the radiations of myelinated fibres, are the essential conditions of the structure of the prosencephalon. In all vertebrates, except the fishes and amphibia, the ganglionic nidi are placed upon the floor of the paraceles. They there form rounded prominences,[p. 128] from which myelinated fibres radiate out towards the periphery of the hemispheres.
The lowermost stratum of the floor of the paraceles is, therefore, composed of the continuations of the ascending and diverging crura. Upon these rest, first of all, the thalami. New tracts of alba issue from the thalami, and join and reinforce the crural bundles that run forwards and outwards below them. These terminal radiations of the crus, at the anterior and external border
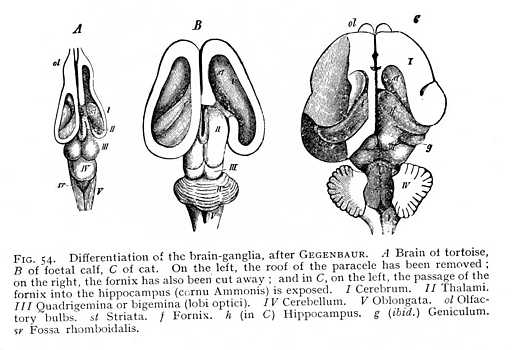
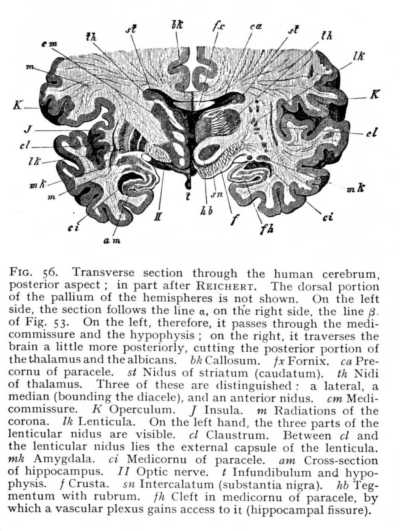
of each thalamus, are, again, intermixed with large ganglionic nidi. The result is that the floor of the paracele rises in a rounded prominence of considerable extent, which forms the anterior and exterior boundary of the thalamus. It is termed the striatum (st Figg. 54, 55). The club-like extremity, lying anteriorly of the thalamus, is called the caput; the narrower portion, surrounding the thalamus exteriorly, the cauda. Striatum and thalamus together cover the entire floor of the paracele. The surface of the striatum is invested with a tolerably thick layer of cinerea, whereas the thalamus (i.e., that portion of its surface which projects into the paracele) is covered by a lamina of alba. Along the border of thalamus and striatum lies a narrow hand of alba, the tenia (stria terminalis, strin cornea: sc Fig. 55). The ganglionic nidi of the striatum appear in the mammalian brain, as three characteristically shaped masses. The first is directly connected with the grey investment of the striatum; it follows the arch of the peripheral surface, and so acquires a curving [p. 129] form, which has given it the name of caudatum (nucleus caudatus: st Fig, 56). It constitutes, with the myelinated masses that begin their course below it, the striatum in the narrower sense. A second and very considerable nidus, the lenticula (nucleus lentiformis: lk), lies to the outside of the caudatum. In vertical section it appears as a triangle, whose apex points towards the internal edge of the striatum, while its base extends far out into the alba of the hemisphere. The lenticular
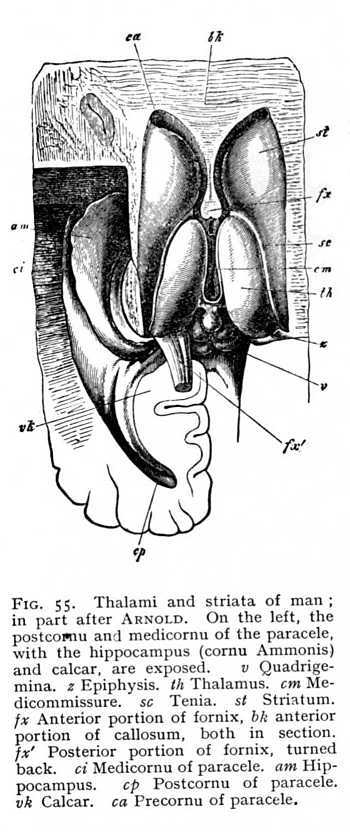
cinerea is divided up by intervening myelinated fibres into three groups, two external and ribbon-shaped, and one internal and triangular. The third and last nidus of the striatum lies outward from the lenticula. It, too, has the form of a narrow ribbon of tissue, and surrounds the third subdivision of the lenticula. It is named, from the closeness of its approach to the brain surface, the rampart or claustrum (nucleus taeniaeformis: cl). Ventrally from the claustrum, and near the cortex of the brain-base, lies yet another small nidus, the amygdala (mk).[6] These ganglionic nidi of the hemispheres take up many of the myelinated fibres that spring from the quadrigemina and thalami; others pass under the striatum and forwards, without coming into connexion with its cinerea. Above the nidi, the myelinated bundles coming up from below radiate out from the whole extent of the striatum, in the most various directions, toward the cerebral cortex. The terminal division of the great longitudinal fibre-tract, that begins in the columns of [p. 130] the myel, then passes over into the columns of the oblongata, and thereafter takes its place among the bundles of the crura, is the corona (corona radiata; m). The factors which most largely determine the arrangement of its fibres are those discussed just now, as concerned in the formation of the paraceles. Since the vascular plexuses that find their way into the cavities spread over the entire floor, the coronal fibres which are to continue the crura cortexward must curve out around the vessels at the periphery, in order to attain their goal.
The terminal division of the prosencephalon consists of the two olfactory bulbs or olfactory gyres (rhinencephalon). In most fishes, the rhinencephalonis so strongly developed that it not seldom surpasses in extent all the remaining portions of the prosencephalon. In the higher classes of vertebrates, and especially in the birds, it decreases in importance; but in the lower mammals it appears again as a structure of relatively considerable size (cf. Figg. 37, 38, 49, 54). In the mammalian brain it forms special gyri, which issue from the brain-base and project to a greater or less degree beyond the frontal portion of the prosencephalon. The olfactory bulbs contain cavities, the rhinoceles, which communicate with the prosocele (paraceles). In some of the mammalian orders, viz., in the cetacea and (to a less extent) in the apes and in man, the rhinencephalon degenerates.[p. 131] The olfactory bulbs lie far back under the frontal regions of the hemispheres and are connected by a narrow stalk, the olfactory tract, to the middle part of the brain-base (Fig. 52, p. 125). The area which serves as point of departure for the tract, the olfactory area, presents a sieve-like appearance due to the incoming of numerous small vessels, and is consequently termed the precribrum (lamina perforata anterior: sp Figg. 47, 52).
The fuller development of the prosencephalon brings with it a radical transformation of the two lateral ventricles, the paraceles. This is due, in part, to the growth of the hemicerebral masses which enclose them, but in part also to the appearance of special
structures which project into the cavities. As the hemispheric vesicle of the mammalian brain arches back over the diencephalon and mesencephalon, the portion that lies behind the Sylvian fossa takes a downward turn (Figg. 36, 42, p. 108, 112). The result is that the paracele possesses two branches, or cornua, as they are termed: a precornu, bounded on the outside by the arched wall of the hemisphere, and a medicornu (cornu inferius) whose extremity is drawn out to a point. The growth of the hemispheric vesicle over the caudex is accompanied throughout its progress (as we have already seen: p. 114 above) by a parallel growth of the aula (foramen of Monro), the original means of communication between prosocele and diacele. As the aula, then, curves over the caudex, at first posteriorly and then ventrally, what was originally its dorsal extremity coincides with the pointed end of the medicornu. The part of the aperture that now lies in the anterior wall of the medicornu forms a fissure (the hippocampal fissure, of which more presently), which is occluded by a vascular plexus from the pia (fh Fig. 56). In fine, therefore, the primitive aula remains open at beginning and end, but is closed over its middle portion by myelinated fibres. These belong to the fornix and callosum, structures which we shall discuss in the following section.
In the brain of the primates (the apes and man), the conformation of the paraceles undergoes yet another change, due to the large development of the occipital portion of the hemispheres. The outer wall of each paracele pushes vigorously backwards before it takes the curve downwards, so that the cavity itself is prolonged in the same direction. We thus have a postcornu (cp Fig. 51, p. 123) in addition to the precornu and medicornu. The backward growth of the prosencephalon stops, as it were, with a jerk, to continue forwards and downwards. This fact is attested both by the outward appearance of the occipital region, and by the shape of the postcornu, which is drawn out into an even finer point than the medicornu. In the apes, the postcornu is smaller than it is in man; in other mammals with strongly developed hemispheres, as e.g. the cetacea, it is no more than a trace or rudiment of what it is later to become.[p. 132]
(b) -- Fornix and Commissural System
At the anterior extremity of the primitive aula, the two hemispheres grow together in the middle line. The resulting strip of alba is termed the terma (lamina terminalis: bd Fig. 43, p. 113). The backward curvature of the hemispheres round the transverse axis of the diencephalon naturally brings with it a corresponding curvature of the terma. Its most ventral and anterior extremity becomes a band of cross-fibres, connecting the two hemispheres, and known as the precommissure (k Fig. 43). In its further course it divides into two lateral halves running longitudinally from before backwards, on either side of the median fissure. We find the first beginnings of these longitudinal fibre-tracts in the birds, but they do not attain to any high degree of development until we reach the mammals, where they constitute the fornix. Closely approximated anteriorly, the columns of the fornix diverge as they pass backwards. The myelinated fibres of their anterior extremity extend ventrally to the brain-base, where they stand in connexion with the alba of the albicantia (Fig. 53, p. 127). The fibres of their posterior extremity are distributed in man and the apes into two bundles, the smaller of which comes to lie upon the inner wall of the postcornu, and the larger upon the inner wall of the medicornu of the paracele. The projection thus occasioned in the wall of the postcornu is termed the calcar (pes hippocampi minor), that in the medicornu, the hippocampus (pes hippocampi major: Fig. 55). These prominences are, however, constituted in part of other factors, which we shall discuss later. In the other mammals, which have not developed a postcornu, and which therefore cannot possess the calcar, the whole mass of fornix-fibres passes over into the hippocampus.[7]
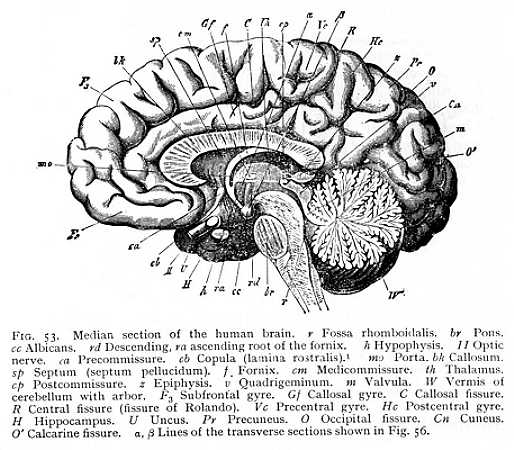
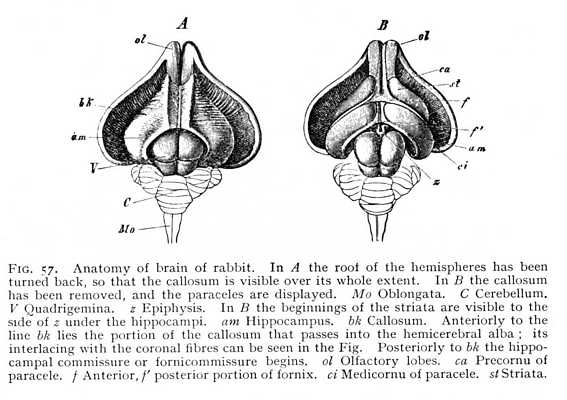
The formation of the fornix appears to stand in intimate relation to that of another transverse fibre-system whose appearance is even more definitely characteristic of the mammalian brain. In the monotremes and marsupials, new fibre-tracts are observed to issue from the hippocampus (cornu ammonis). They run dorsally of the incoming fornix-fibres, and pass above the diencephalon to the opposite half of the brain, where they terminate, as they began, in the hippocampus. The transverse commissure that thus arises between the two hippocampi is the original primule of the callosum. In the non-placental mammals, in which the callosum is this restricted to a mere cross-commissure between the two hippocampi, the precommissure [p. 133] is very strongly developed, as it is in the birds, though a free space is left between it and the callosum. In the placental mammals, the hippocampal commissure is reinforced by additional transverse fibre-tracts, which radiate out into the hemicerebral alba at large. They make their first appearance at the anterior end of the future callosum, so that the development of the callosum itself proceeds from before backwards. At the same time, the precommissure decreases in size, and enters by way of a thin and still transverse lamella of alba (Fig. 53 ca) into connexion with the anterior extremity of the callosum, the 'beak' or rostrum. This junction of precommissure and rostrum results in the anterior occlusion of the intercerebral fissure. Between the broad posterior extremity of the callosum, the splenium, and the dorsal surface of the cerebellum, there still remains, however, a narrow passage, by which the diacele can communicate with the surrounding space (the passage is visible in Fig. 53 as the dark space between epiphysis and splenium). This is continued laterally as a narrow cleft, leading into the paracele. We have, in it the remnant of the original anterior roof-slit, whereby the vascular plexuses gain access to the three anterior brain-cavities (p. 114).
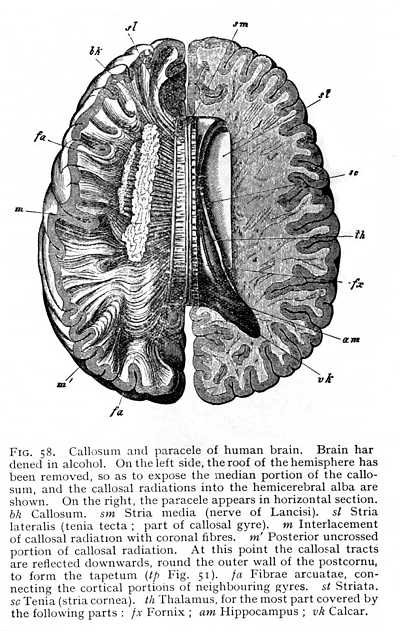
In most mammalian brains, the hippocampal commissure persists as a relatively large portion of the entire callosum (bk Fig. 57 A). Moreover, since the occipital brain is here but little developed, and the anterior brain-ganglia, the thalami and striata, also decrease very considerably in mass, the hippocampus is brought forwards to the point of origin of the fornix. The fornix itself immediately separates on either side into two divisions, the one of which forms the anterior and the other the posterior boundary of the hippocampus (f and f' Fig. 57 B).[8] It is, however, not until we reach the higher mammals that we find any considerable development of the fornix. Between the callosum and the deeper-lying fornicolumns are now spread two thin vertical lamellae of alba, enclosings a narrow cleft-like cavity. These are the septa (septa pellucida: sp Fig. 33). Fornix and septa occlude the internal openings of the paraceles; nothing is left but the beginning of the original aula, just behind the anterior place of origin of the fornicolumns (porta or foramen of Monro: mo Fig. 53; cf. i Fig. 43, p. 113). Between the lateral halves of the septa is the cleft-like cavity just mentioned; it communicates posteriorly with the diacele, and is termed the pseudocele (cavum or ventriculus septi pellucidi). The callosal radiations form the roof and a portion of the outer wall of the paraceles. As external capsule, they skirt the external margin of the lenticula. On their way to the cortex, where they terminate, they interlace at all points (the posterior strands excepted) with the fibres of the corona. The posterior fibres, coming from [p. 134] the hippocampi and their neighbourhood, do not receive any admixture of coronal fibres. In the lower mammals, they appear simply and solely as the hippocampal commissure (Fig. 57 A); in the primates, they divide into two parts; an internal, passing over into hippocampus and calcar (am and vk Fig. 55); and an external, which curves ventrally in front of the coronal fibres running to the cortex of the occipital lobe (m' Fig. 58), and forms the outer wall of the postcornu of the paracele. This is termed the tapetum (tp Fig. 51, p. 123).
We have seen that the fornix is the fibre-tract proceeding from the terma (lamina terminalis) of the aula; and we have followed the course which it takes as the outgrowing hemispheres arch over the brain-caudex. The same direction is taken by the portion
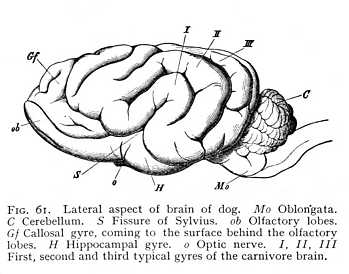
of the hemicerebral wall lying immediately anterior to the terma. There is a difference, however. The floor of the hemispheres is continuous from the first, and the fornix has, therefore, no investment of grey matter. This anterior portion, on the other hand, which comes to lie dorsally to the fornix as the result of the hemispheric curvature, is not included in the original area of continuity, and is accordingly covered with a layer of cinerea over its median surface. After the callosum has forced a passage across the brain, it is separated by the callosal fibres from the fornix, and forms a longitudinal gyre running dorsal to the callosum. It is termed the callosal gyre (gyrus fornicatus or cingulum: Gf Fig. 53, p. 127).[p. 135]
In certain mammalian brains, where the frontal part of the prosencephalon is but little developed, while the callosal gyre is large,
it can be traced anteriorly from a point directly behind the base of the olfactory tract. After curving over the callosum, it again emerges posteriorly at the brain-base. Here it passes over into a gyre lying behind the Sylvian and bounding the median fissure; the hippocampal gyre, the outer wall of the hippocampus (H Fig. 53). Where the callosum begins, the layer of cinerea ceases; the lower surface of the callosal gyre, the surface adjacent to the callosum, consists of unmixed white matter. The sole exception is a narrow stria of cinerea, isolated from the rest of the cortex, which has persisted in its posterior portion. This is known as the fasciola (fc Fig. 59); it lies immediately above the callosum. The fasciola is free, over its whole extent, from any admixture of alba; so that the longitudinal myelinated fibres upon which it rests are entirely separated from the remaining white matter of the callosal [p. 136] gyre. When the gyre is removed from the callosum, these fibres, together with the fasciola which invests their posterior portion, remain attached to it, they appear as a myelinated stria, and have received a special name, tenia tecta or lateral stria (sl Figg. 58, 59). The importance of this separation of lateral stria, and fasciola from the rest of the myelinated and cortical substance of the callosal gyre lies in the fact that the structures remain isolated at the point of transition from callosal to hippocampal gyre.[9] Alba and cortex of the callosal gyre pass directly over into alba and cortex of the hippocampal. Really, therefore, the two are but one: the only difference between the parts being that the callosal gyre is not invested with cinerea over its ventral surface, the surface adjacent to the callosum, whereas with transition to the hippocampal gyre the cortex spreads out again over the entire surface of the convolution. Now at the point where the callosal gyre leaves the splenium and becomes the hippocampal gyre, -- at the point i.e. where the cortex which has previously invested the inner surface only extends over the ventral as well, -- the lateral stria divides from the rest of the white matter of the gyre, and appears upon the surface of the gyrus hippocampi. This means, of course, that the fasciola, which lies just below the lateral stria, must divide from the rest of the cortex; the lateral stria forms a partition between cinerea and cinerea. The result is that we have, at the point in question, a cortical layer covered by a lamella
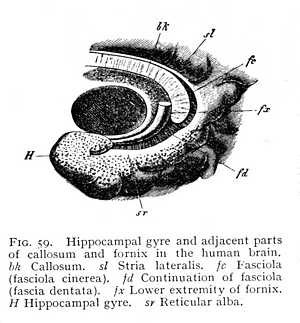
of alba, and this again covered by a grey cortex. The two most superficial layers, lateral stria and fasciola, are, it must be remembered, strictly limited in area; they extend only over the hippocampal gyre. Indeed, they cover only a portion of that; for the white and grey areas are not coincident. The alba of the lateral stria is distributed over the entire cortex of the hippocampal gyre, as an extremely thin reticular layer of white fibres. This reticular alba is the only white layer that appears upon the cortical surface of the hemispheres (sr Fig. 59; cf. also H Fig. 53, p. 127). The fas-[p. 137]ciola, on the other hand, retains its ribbon-like form; it covers, not the whole radiation of the white fibres of the lateral stria, but only a certain group of them, the group lying in the fissure which forms the interior boundary of the hippocampal
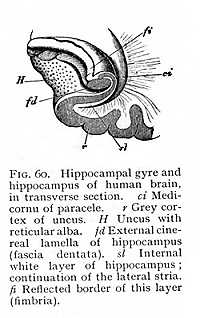
gyre. From the peculiar toothed appearance that it has at this part of its course it is known as the fascia dentata (fd Fig. 59). The fissure which forms the interior boundary of the hippocampal gyre corresponds to the hippocampal projection into the medi-cornu of the paracele. The formation of the hippocampus -- to which, as we have seen, fibres from callosum and fornix contribute -- is thus completed by contributions from the various regions of the callosal gyre. The white layer which invests the paracele-surface of the hippocampus is formed by the fibres of callosum and fornix (Fig. 60). It is followed by a first cinereal layer, the cortex of the hippocampal gyre (r); externally to that comes a second layer of alba, the continuation of the lateral stria or the reticular alba (H) distributed over the cortex of the gyrus hippocampi; and, lastly, beyond that follows a second cinereal layer, the fascia dentata, the continuation of the fasciola (fd). This extends, as we have said, only to the fissure which corresponds to the hippocampal projection. The same fissure forms the inner boundary of the reticular alba. Along the line at which the alba ceases, the grey matter of the fascia dentata is continuous with the cortex of the hippocampal gyre; so that here the two cinereal layers which fill the interior of the hippocampus are brought into contact. At the precise point where this transition takes place, the internal white investment of the hippocampus terminates in a free reflected border, the fimbria (fi).
(c) -- The Development of the Outward Conformation of the Brain
We have now passed in review those divisions of the brain which appear in the general course of neural evolution.
This articulation into parts is paralleled by a series of changes in outward form, the final outcome of which is dependent partly upon the degree of general development to which the particular brain has attained, and partly upon the relative growth of the individual parts. In the lowest vertebrates, the brain has gained but little upon the simplest embryonic form which is given with the separation [p. 138] of the primitive brain-vesicle into its five subdivisions. The whole range of structural difference is here practically exhausted by differences in the relative size of these subdivisions; the only further determinant of final brain-form is the development of the olfactory lobe as an outgrowth from the prosencephalon. A much greater variety of configuration appears as soon as the pallial structures begin to invest the brain-caudex. The covering of quadrigemina (bigemina) and cerebellum by cerebral hemispheres and of oblongata by cerebellum, and the degree of encephalic flexure, bring in their train a long series of structural peculiarities; and the list is still further swelled by differences in the outward form of the hemispheres, by the development or lack of development of the cerebellar pilea, by the corresponding appearance or non-appearance of certain nidal structures (such as the olives) on the oblongata, and by the development of a pons.
The point where the cerebral hemisphere originally rested upon the brain-caudex is marked, in all mammalian brains without exception, by the Sylvian fossa (S Fig. 42, p. 112). In the higher mammals, the edges of the fossa draw together, so that we
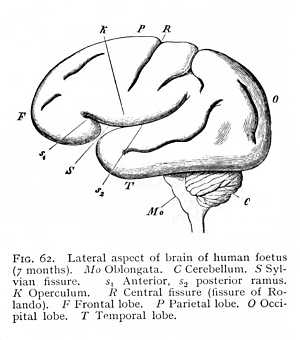
find in its place a deep fissure, the Sylvian fissure (fissura Sylvii). The fissure usually runs obliquely, from posterior-dorsal to anterior-ventral; its divergence from the vertical is determined by the growth of the occipital brain and its extension over the posterior parts of the system (Fig. 61). In the highest mammalian order, that of the primates, the Sylvian fissure undergoes a final and characteristic transformation. The frontal and occipital brains here develope simultaneously and the fossa formed by the growth of the hemispheres over the caudex consequently appears, at the very beginning of the embryonic life, as a roughly outlined triangle, lying base upward. The edges, above, below and behind, then grow towards each other, and the fossa closes to form a Y-shaped fissure (S Fig. 62), dividing into an anterior and a posterior ramus (s1 and s2. Cf. also Fig. 65) The part of the hemisphere that lies between the two rami, and roofs in the original fossa from above, is termed the [p. 139] operculum (K). If the operculum is turned back, and the floor of the Sylvian fossa exposed, the underlying hemispheric surface proves to be bulged out and, like all the rest, divided by fissures into a number of gyres. The brain-region which is concealed and isolated in this peculiar way is known as the central lobe or island (insula Reilii; Fig. 56 J, p. 130). The two rami of the Sylvian fissure form the customary points of departure for the division of the hemicerebra of the primate brain into separate regions or lobes. The portion lying anteriorly of the anterior ramus is termed the frontal lobe (F Fig. 62); the space included between the rami, the parietal lobe (P); the region behind the Sylvian fissure, the occipital lobe (O); and the area situated ventrally of it, the temporal lobe (T). These lobes pass into one another, on the convex surface of the brain, without any sharp line of demarcation.
Not only does the Sylvian fissure divide the surface of the hemispheres at large into a number of lobes: there are certain other and smaller areas that are marked off from their surroundings by furrows or fissures. Thus the longitudinal fibre-tract running dorsal to the callosum from before backwards, and then curving ventrally round the splenium, -- the area with which we have become familiar as the callosal gyre, -- may be recognised by the presence of definite fissures, separating it from the surrounding parts (Gf Fig. 53, p. 127). In all mammalian brains, in particular, we can trace on the median surface of the hemispheres the margin along which the investment of the inner portion of the callosal gyre is deflected into the medicornu of the paracele (hippocampal fissure: fh Fig. 56, p. 130); while in most of them the callosal gyre is also bounded during its course upwards over the callosum, by a longitudinal fissure, the callosal fissure (sulcus callosomarginalis: C Fig. 53). In the same way, the olfactory lobe or olfactory gyre at the base of the prosencephalon is almost always set off by an inner and an outer fissure, the entorhinal and ectorhinal fissures; though in the human brain the two have fused to one (sr Fig. 52, p. 125). [p. 140] All these fissures and furrows are occasioned, then, by the growth of the hemispheres round their point of application to the diencephalon (Sylvian fissure), by the occlusion of the external fissure of the medicornu (hippocampal fissure), or by the course of determinate bundles of myelinated fibres appearing on the ventral and median surface of the hemispheres (callosal, entorhinal and ectorhinal fissures). Since the structural relations that condition them are characteristic of the mammals as a class, they form, as soon as they can be traced at all, entirely constant features of the mammalian brain.
But there are other fissures, less uniform in their course, which give the brain-pallium of the mammals a variously convoluted appearance. The surface of cerebrum and cerebellum is split up by them into numerous gyres. The cerebellar gyres are, on the whole, arranged with more regularity than the cerebral; they form narrow ridges, set vertically upon the underlying alba, and following for the most part a transverse direction. On the cerebrum, whose folds are not unlike the convolutions of the intestine, it is often difficult to recognise any definite law of gyre-formation. The common cause of all these ridges and foldings of the brain-surface is evidently to be found in the disproportionate growth of the cortex and of the myelinated tracts that radiate into it. When a body increases in mass, its surface, of course, enlarges less rapidly than its volume. But the cells of the brain surface have to take up the fibres of the interior alba; and so surface-extent must be roughly proportional to volume, and this relation must be maintained with approximate constancy throughout the whole period of development. It is obvious, then, that the cortex has no way of keeping pace with the increase of alba except by folding. And it is for this reason that, both in the organic series and in the course of individual development, the convolution of the brain-surface increases with increase of the size of the brain.
The convolution of the cerebellum is found in its simplest form in the birds. The avian cerebellum has no pilea, and so appears, in dorsal aspect, as an unpaired structure of a more or less spherical or ovoid form. The surface of this organ us split up into transverse folds, roughly circular or elliptical in shape, all intersecting in an axis laid transversely through the centre of the sphere or ovoid: the transverse axis is, therefore, in this case the common axis of convolution for all gyres visible upon the surface of the cerebellum (Fig. 49 A, p. 121). If we bisect the organ at right angles to the direction of this axis, we find that the depth of the fissures bounding the separate prominences varies: the ridges fall into groups of two or three, marked off from one another by shallow depressions, but divided from the neighbouring groups by deeper fissures (Fig. 40 B, p. 110). In the mammals, the convolution becomes more complicated; each of the groups of ridge-like prominences marked off by the deeper fissures contains a large number of [p. 141] separate gyres. Moreover, it frequently happens that several of these groups are isolated by dividing fissures, and so form still larger units, lobes. The consequence is that most of the gyres come to lie in the depth of the larger fissures, and that only the terminal lamellae appear on the surface. Hence we have in sections the well known representation of a tree, with its spreading leaves and branches, termed by the older anatomists the arbor vitae (av Fig. 50, p. 121; cf. also W Fig. 53, p. 127) Further, while this fissural differentiation is in progress, the median portion or vermis of the cerebellum is reinforced by the large, bilaterally symmetrical pilea. If the arrangement of gyres upon the pilea is fairly regular, as in man, the principal axis of convolution is again the transverse. The rule is broken, however, along the anterior and posterior margins, where the gyres gradually change their course to take an oblique or even longitudinal direction, all alike converging towards the point of attachment of pileum to vermis (Fig. 50). For the rest, many mammalian cerebella evince a great variety in the course of their gyres, more especially upon the pilear surfaces, so that no definite law of gyre-formation can be made out. This is most apt to occur in brains whose cerebellum is richly convoluted. In the human cerebellum itself there are certain divisions, isolated by important fissures,[10] in which the course of the gyres diverges more or less widely from the general trend.
An increase by convolution of the superficial extent of the cerebrum is found only in the higher vertebrate classes. And the brains even of the lowest orders of mammals show, at most, only the fissures and gyres described above (Sylvian fissure, hippocampal fissure, etc.), which are due to special causes, distinct from those that produce the remaining ridges and furrows. When once these latter have appeared, however, they persist in practically the same pattern throughout the mammalian series, up to and including the primates. The rule is that all fissures and gyres running from before backwards take a course which is approximately parallel to that of the median fissure; most of them are also reflected in a curve round the Sylvian fissure (cf. Fig. 61 I, II, III, p. 138). That is to say, the gyres take the same direction that the hemispheres take in their growth round the brain-caudex, -- the direction from before backwards; and they are reflected round the point of application to the diencephalon in a curve which repeats the curve of hemicerebral growth. The degree of curvature is determined by the depth and extent of the Sylvian fossa or fissure. The number of longitudinal folds which thus appear on the surface of the cerebrum varies in general, in the various mammalian orders, between two and five. In many cases, one or another of them forms a junction, at some [p. 142] point of its course, with the neighbouring fold; and very frequently less well marked, secondary folds are formed, making an angle with the primary. The convolution thus appears as an irregularly meandering system, in which the general law of direction is more or less obscured. The case is very different with the formation of folds in the frontal region of the cerebrum. A little anteriorly to the Sylvian fissure, the longitudinal gyre-tract is replaced, suddenly or gradually, by an approximately transverse ridging. At the same time, the secondary cross-fissures are often disposed radially about the Sylvian fissure. In accounting for this mode of fissure-formation in the frontal brain-region, we must remember that in all mammals, except the cetacea and primates (the orders in which the olfactory gyres are to some extent degenerate), the callosa1 gyre comes to the surface in the frontal area, and is separted from the more posterior gyres, at its point of issue, by a transverse or oblique fissure; anteriorly, it passes directly over into the olfactory gyre, from which it is also separated, though as a rule less distinctly, by another cross-fissure (Fig. 61, Gf). The point at which the callosal gyre comes to the surface lies in some instances at a very short distance from the anterior boundary of the brain. This is the case e.g. in the carnivores, where the gyre is exceedingly broad, so that it and the olfactory gyre together occupy the entire area elsewhere taken up by the frontal brain. In other brains, the point of issue lies more posteriorly. The exposed portion of the callosal gyre is then, as a general rule, longer than it is broad; it merely fills a narrow space to either side of the anterior part of the intercerebral fissure. The cross-formation of which we are speaking is not confined, however, to the folds caused by the protrusion of the callosal and olfactory gyres; all the other fissures of the anterior brain-region take the same transverse direction. In some cases, the folds that run longitudinally in the occipital area are deflected anteriorly into a transverse course; in others, the longitudinal furrows are suddenly interrupted, and replaced by cross-fissures. We have a salient instance of the former arrangement in the brain of the carnivores (Fig. 61), which is characterised by the regularity and symmetry of its convolution; while the second type is represented by most of the other richly convoluted mammalian brains, though here, too, certain of the longitudinal fissures are not infrequently continued in the transverse direction. There are, as a rule, two principal fissures of this and crossing the frontal brain, and the number holds, whether the fissures are completely independent or pass over posteriorly into longitudinal fissures. To these must be added, further, the posterior limiting fissure of the callosal gyre, and the fissure separating the callosal and olfactory gyres. There is thus, upon the average, a total of four transverse fissures in the frontal brain-region.[11][p. 143]
All these folds, longitudinal and transverse alike, are ordinarily visible only on the upper and outer surface of the hemispheres.
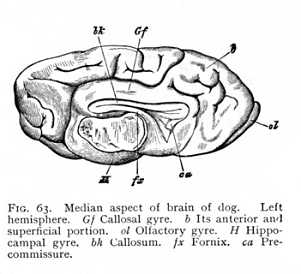
The base of the brain is, in most instances, entirely occupied by the gyres and fissures previously described, -- anteriorly, i.e., by the olfactory gyre, and posteriorly by the hippocampal gyre (ob, H Fig. 61). If anything else can be seen, it is at most simply a narrow margin belonging to the outermost gyres of the brain-surface. Again, the greater number of brains, when viewed in median section, show only the callosal gyre and its continuations, posteriorly into the hippocampal gyre, and anteriorly into the olfactory gyre (Fig. 63); though in certain cases, where these structures are not prominent -- as in the brains of the cetacea, of the apes and of man -- portions of the superficial gyre-tracts may also make their appearance. But these brains deviate markedly in other respects as well from the general law of fissuration evinced by the mammalian brain. In the cetacea, whose organs of smell, peripheral and central, are completely atrophied, the callosal gyre does not come to the surface, and no olfactory gyre is present at all. The brain is extraordinarily broad; and the principal superficial fissures run over its whole length from before backwards, as they do in the occipital brain-region of the other mammals.
In the primates, the fissuration of the brain follows a special law of evolution, characteristic of the order. The olfactory gyre, which has dwindled into the olfactory bulb, lies concealed upon the brain-base. The callosal gyre appears on the surface, but only in the occipital, not in the frontal region of the brain. As the gyre curves posteriorly round the splenium, on its passage into the uncus, it sends off a branch to the surface of the occipital lobe. This divides into two lobules, the cuneus and precuneus (Pr, Cn Fig. 64). The intruding area forms a sort of island, bounded before and behind by other gyres, from which it is ordinarily divided by fissures. The cuneus and precuneus are also themselves separated by a deep transverse fissure, the occipital fissure (O). The same transverse arrangement of gyres and fissures obtains, further, over the entire occipital portion of the brain, from
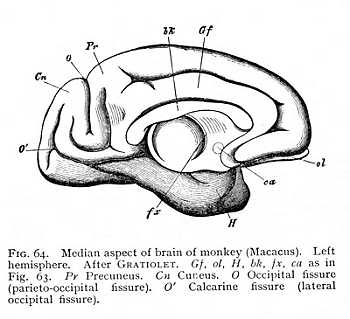
the anterior extremity of the main ramus of the [p. 144] Sylvian fissure (the Sylvian fissure in the narrower sense) to the extreme posterior limit of the occipital lobe. Anteriorly, the principal fissure which runs in the transverse direction, from above downwards, is the central fissure (fissure of Rolando: R Fig. 65). It is bounded before and behind, in the brain of man and of the higher apes, by two transverse folds, the precentral and postcentral gyres (VC, HC Fig. 65). These are separated from the adjacent parts -- the precentral from the frontal gyres, and the postcentral gyre from the precuneus -- by shorter cross-fissures. Finally, a deep transverse fissure is found at the posterior limit of the occipital brain. This is the calcarine fissure, which separates the cuneus from the gyres extending downward to the brain-base (O'). We have, then, in all, on the surface of the occipital brain, five well-marked transverse fissures, three of which belong to the offshoots of the callosal gyre and the adjoining parts. On the other hand, the fissures and gyres of the frontal and temporal areas -- i.e. of the region anterior to the presylvian and ventral to the subsylvian fissures -- tend in general to pursue a longitudinal course, during which they are necessarily reflected in a curve round the main ramus of the Sylvian fissure. Three of these longitudinal folds can be distinguished in the frontal, and three in the temporal region; they constitute the three frontal and temporal gyres (F1-F3, T1-T3), all of which run far enough to be visible at the base of the brain (Fig. 52, p. 125). The three temporal are adjoined posteriorly to the three occipital gyres (O1-O3). In the area of transition from the occipital to the temporal region, the folds follow a course that lies midway between the transverse and the longitudinal; so that the three parietal gyres (P1-P3) show a gradual passage from the one to the other direction. There is no parallel to this in the frontal region: the three frontal gyres are interrupted suddenly and at right angles by the downward course of the precentral. The essential difference, then, between the brain of the primates [p. 145] and that of the other mammals lies in the fact that the primates have the transverse fissures in the occipital, and the longitudinal fissures in the frontal region, whereas most of the other mammals have these relations exactly reversed. There is a corresponding difference in the course of the callosal gyre. In the primates, the callosal gyre comes to the surface in the posterior, in the lower mammals in the anterior region of the brain: a fact which can be brought out most clearly by the comparison of a primate brain and the brain of another mammal in median section (Figg. 63, 64). These differences are probably referable to the divergence of the laws of growth in the two brain-forms. In most mammals, the developing brain has a strong lateral growth in the occipital region, while the frontal area remains narrow; so that the whole organ is wedge shaped, tapering from behind forwards. In the brain of the primates, on the contrary, the growth of the occipital
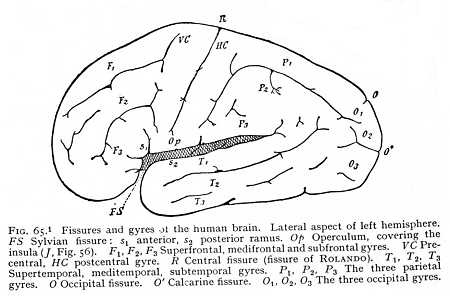
region is predominantly in the longitudinal, and the growth of the frontal region predominantly in the lateral direction. The organ thus assumes the form of an ovoid, whose lateral halves are closely apposed anteriorly, but posteriorly gape apart; while the lesser height of the posterior portion leaves room for the growth of the cerebellum beneath it. Embryology tells us that the cross-fissures upon the cerebrum of man (and probably of the primates at large) represent the original foldings of the cortex.[p. 146] According to ECKER, they begin to form upon the smooth cerebral surface as early as the fifth month, whereas the first traces of longitudinal fissuration make their appearance in the course of the seventh month.[12] Four or five of these transverse fissures, disposed radially with more or less of regularity about the Sylvian fissure, may be traced upon the foetal brain. The most strongly marked among them becomes the central fissure. In the apes, the central fissure is less prominent than in man; but this defect is compensated by the greater development of the more posteriorly situated occipital fissure, which has accordingly been termed the ape-cleft (Affenspalte).[13] The calcarine fissure, lying still more posteriorly, can hardly been seen at all on the human brain except in median section (Fig. 53, p. 127, and O', Fig. 65). It is this fissure whose projection into the postcornu of the paracele forms the calcar of the primate brain (vk Fig. 55, p. 129). In man, it joins the occipital fissure at an acute angle, so that the cuneus shows as a wedge-shaped lobule, apparently isolated from the callosal gyre (Cn Fig 53). In the apes, the calcarine fissure is shallower, and the connexion of the cuneus with the callosal gyre can accordingly be seen at once (Fig. 64). But while the portion of the primate brain that lies behind the central fissure thus developes several well marked cross-fissures, the transverse fissuration of the anterior half is much less pronounced. In place of it, we find throughout the frontal and temporal regions the longitudinal fissures and gyres which we have mentioned as appearing at a later stage of embryonic development. The three longitudinal folds that are distinguishable upon every primate brain give rise to the three frontal (super-, medi-, sub-) and the three temporal (super-, medi-, sub-) gyre-tracts (Fig. 65). These, however, do not form (as they do in many of the other mammals) a, continuous convolution, arching over the Sylvian fissure. On the one hand, the three frontal gyres are interrupted by the precentral gyre. On the other, only one of the temporal gyres (the supertemporal) runs a complete course to the postcentral gyre, curving boldly upwards round main ramus of the Sylvian fissure; the other two are deflected by the precuneus and cuneus, the two lobules bounded by the other radial fissures of the occipital brain, and then continued on the parietal brain-surface into the three parietal gyres (P1-P3). These latter, situated as they are at the meeting point of different gyre systems, show the least regular course of all: the third parietal gyre (P3), which bounds the posterior extremity of the Sylvian fissure, has been named from its shape and position the angular gyre. On the median surface of the brain, the first and second parietal gyres form the boundary of the precuneus and cuneus (Pr, Cn, Fig. 53, p. 127) which here appear as direct continuations of the callosal gyre (Gf). On the brain-base, the subtemporal gyre is connected anteriorly with the club-shaped termination of the hippocampal gyre; posteriorly, it is continuous with the external ramus of an U-shaped gyre-tract which occupies the base of the occipital brain and whose inner ramus is lost in the stem of the hippocampal gyre (O Fig. 52, p. 125). The anterior portion of the brain-base is occupied by the ventral deflections of the three frontal gyres. The medifrontal and subfrontal become continuous on the margin of the Sylvian fissure (F2, F3, Fig. 52)[14][p. 147]
The law of fissuration of the brain-surface is, in the author's opinion, the resultant effect of two sets of causes: of the tensions produced by the growth of the brain itself, and of the influence upon the brain of the enclosing skull- capsule. The former condition may be appealed to in explanation of the fissures that appear at the earliest period of development. If a surface is to increase its extent by fold-formation, its ridges will of necessity follow the direction of least resistance. If the surface is more tensely stretched in the transverse than in the longitudinal direction, therefore, it will fall into transverse folds, or ridge up about a transverse axis, just like a damp piece of paper that one holds in the hands and pulls out to right and left. If, on the other hand, the tension is greater in the longitudinal direction, then the folds will be longitudinal, the ridges disposed about a longitudinal axis. Suppose, then, that the fissuration is uniform, all the folds takings a single direction: this will mean that the difference of surface-tension remained constant over the whole period of growth. Irregularity of folding will mean, on the contrary, that the line of greatest tension has varied. Now if a structure grows in different directions with varying rapidity, its surface-tensions must vary in these same directions; and the direction of greatest tension must lie at right angles to the direction of greatest energy of growth. A growing structure, i.e., may be regarded as a coherent elastic body, in which the deformation caused by growth at any given point increases the tension at all other points, the tension reaching its maximum in the parts where the intrinsic deformation is minimal. The fissuration of the cerebellum, where the laws of growth and folding are comparatively simple, seems to confirm this principle; and the confirmation is all the more striking, since the position of the organ may be supposed to exempt it from the influences of the skull-form. The growth Of the cerebellum, during the entire period of its development, is predominantly in the longitudinal direction. Its greatest surface-tension must, therefore, lie at right angles, in the transverse direction; and this is, as a matter of fact, the direction of its fissuration. We should expect, on the same principle, that the fold-formation of the primate cerebrum would show two stages, corresponding to two distinct periods of growth: a first, in which the direction of maximal growth is the same for the whole brain, from before backwards; and a second, in which the energy of growth over the frontal and temporal regions is greater in the transverse direction. As a matter of fact, a comparison of embryonic brains at different stages of development shows, at the first glance, that the ratio of the two principal diameters of the human brain changes very considerably during the period of structural elaboration. In the first weeks of development, the whole brain is approximately spherical in form; the longitudinal diameter is but little larger than the greatest transverse diameter. The line of greatest breadth crosses the brain behind the Sylvian fissure, or, more correctly, since the temporal lobes are not yet developed, behind the Sylvian fossa. As the fossa closes to form the fissure, the greatest transverse diameter is shifted anteriorly, crossing the brain at the point where the temporal lobes grow over the margin of the fissure. During this whole period, however, the longitudinal diameter of the hemispheres is gaining more and more upon the cross-diameter; so that as early as the third month the ratio of the two is 1: 0.9, while in the course of the fifth and sixth it has fallen to 1: 0.7. This is the time of formation of the first permanent fissures, all of which, without exception, take the transverse direction.[p. 148]
During the fifth month, the central, occipital, and calcarine fissures make their appearance; and in the course of the sixth, the other primary radial fissures are added to their number.[15] From the end of the sixth month, the laws of brain-growth begin to change. The total form of the brain, as expressed in the ratio of the longitudinal to the greatest transverse diameter, remains practically the same; but the growth of the various parts differs widely from the rule that has previously obtained. If we compare the dorsal aspects of foetal brains, from the sixth to the seventh month, we remark at once that the region extending posteriorly from the central fissure is increasing in length and breadth in approximately the same ratio, but that the frontal portion of the brain is growing more strongly in the transverse than in the longitudinal direction. The temporal lobe undergoes a similar modification. Even in the foetus of six months, its anterior extremity extends almost as far as the ventrally deflected margin of the frontal lobe; but it is still narrow, so that the Sylvian fossa lies wide open. In the following months, the growth of the temporal lobe is most vigorous in the upward, and comparatively slight in the longitudinal direction, and, as a result, the fossa closes to form the fissure. Now the changes here sketched are exactly coincident with the formation of the second system of folds, the longitudinal fissures. Since the growth of the frontal brain is predominantly a transverse growth, the frontal gyres must take a predominantly longitudinal direction. And since the most rapid growth of the temporal lobes is upwards, the folds formed upon them must again run from behind forwards, taking a curved path round the Sylvian fissure. Not only do the newly formed fissures follow this direction, over both regions of the brain surface, but some of those that were originally disposed radially about the Sylvian fissure have their course changed to the longitudinal or arching type. Thus the central fissure itself assumes an oblique position; and the subfrontal and supertemporal fissures, whose primules in the sixth month are radial or transverse, have their direction altered, and take their places in the longitudinal fissure system. The case is different with the portion of the brain-surface that lies between the central fissure and the posterior margin of the occipital lobe. Here the transverse fissures, while they increase in depth and extent, for the most part retain their original positions, and only gradually, as they approach the temporal lobe, pass over into a longitudinal path.[16]
The resistance of the skull-capsule is, necessarily, antagonistic to brain-growth. It is probable, however, that this influence is exerted only in the latest period of the embryonic life, and for a time after birth, before the skull has acquired its permanent form. Its principal condition is the varying rapidity of bone-growth along the various sutures, and the successive closure of these as growth advances. If the growing brain meets with an external resistance such as would be offered by the skull, it must evidently fold upon itself; and the direction of the folds will be the direction of least resistance. Where the skull is of the dolichocephalic form, the course of the fissures will thus be predominantly longitudinal, and where it is brachycephaiic, predominantly transverse in direction. As a matter of fact, a connexion of this kind between skull-form and domi-[p. 149]nant gyre-trend has been made out by L. MEYER [17] and RÜDINGER.[18] But the actual convolution of a given brain will, of course, always be the resultant of the two sets of conditions: the intrinsic tension; of growth, and the extrinsic resistances. The former will manifest themselves most clearly in the course of the original fissures; the latter in the modifications superinduced upon the original convolution at a later stage of development.
Notes
[1] The 'ventricles' are counted from before backwards, with the mesocele (Sylvian aqueduct) omitted: two lateral ventricles (paraceles), third ventricle (diacele), fourth ventricle (metacele or metepicele).
[2] HIS, Die Formentwickelung des menschlichen Vorderhirns. In Abh. d. k. sächs. Ges. d. Wiss., math.-phys. Cl., xv., 1890, 675.
[3] In birds, the sinus rhomboidalis is not closed by proliferation of the nerve-mass, but remains permanently open. It thus resembles the fossa rhomboidalis, the continuation of the myelocele in the oblongata, which persists as an open depression in all vertebrate forms.
[4] Cf. below, Ch. vii. This 3
[5] These are the oculomotor and the trochlear nerves. The third oculomotor nerve, the fifth cranial or abducent ocular, arises in the most anterior region of the oblongata.
[6] Many anatomists restrict the name striatum to the caudatum alone, i.e., do not extend it to embrace the lenticula, The claustrum and amygdala must be regarded, from the form of their cells, not as true ganglionic nidi, but as parts of the cortical cinerea, from which they are separated by an intercalated lamina of alba.
[7] The question whether the apes have, like man, a postcornu to the paracele and a calcar (pes hippocampi minor) led to a not very profitable controversy between OWEN, who took the negative side, and HUXLEY. Cf. HUXLEY, Evidence as to Man's Place in Nature, 1863, 100, 113. The older writers on the ape-brain figure the postcornu. Cf. e.g. TIEDEMANN, Iconces cerebri, 1825, 54. OWEN himself, in his later work, describes the rudiment of a postcornu in the dolphin (Anatomy of Vertebrates, iii. 10). In the anthropoid apes, as HUXLEY has shown, the calcar exists as well as the postcornu, only at a lower stage of development than in man.
[8] In human anatomy, the portion of the callosum which connects the two hippocampi is termed the psalterium.
[9] The nerves of Lancisi, or striae mediae, belong not to the callosal gyre, but to the callosum itself. See sm Fig. 58.
[10] Here belong more especially the flocculus (fl Fig. 52, p. 125) a small feather-like outgrowth from the posterior margin of the middle peduncle (crus ad pontem), and the amygdala or tonsil (to), an ovoid prominence covering the oblongata between the inferior portion of the vermis and the pileum of either side.
[11] In the first three editions of this work, the statements of the text are illustrated by cuts of a series of mammalian brains. See 3rd ed. Fig. 48, p. 86.
[12] ECKER, Archiv f. Anthropologie, iii., 1868, 203. PANSCH, ibid., 227.
[13] The morphological relations of the Affenspalte (fissura perpendicularis externa; Wilder's pomatic fissure) are, in reality, more complicated than the author here represents them. -- TRANSLATOR.
[14] BISCHOFF, Abhandl. d. bayer. Akad. d. Wiss., x., 1868. ECKER, Die Hirnwindungen des Menschen, 1869. PANSCH, Die Furchen und Wülste am Grosshirn des Menschen, 1879. FLATAU and JACOBSOHN, Handbuch der Anatomie und vergleichenden Anatomie de Centralnervensystems der Säugethiere, i., 1899.
[15] ECKER. Archiv f. Anthropologie, iii., 1868, 212. Cf. the cuts of embryonic brains in the first three editions of this work. 3rd ed., Fig. 52, p. 93.
[16] In the first edition of this work (p. 101), I have cited measurements of embryonic brains, which serve to confirm the statements of the text.
[17] Centralblatt f. d. medicin. Wiss., 1876, No. 43.
[18] RÜDINGER. Ueber d. Unterschiede d. Grosshirnwindungen nach d. Geschlect beim Fötus u. Neugeborenen, 1877, 5 ff.
Footnotes
[1] This figure [Fig. 53], with several others, has been printed without change in all editions of the Physiologische Psychologie. The reader will observe that the line from the abbreviation cb is carried to what has the general position of the terms (lamita terminalis). The real copula (the weisse Bodencommissur of the German; Henle's commissara baseos alba) is neither designated nor even shown in the Fig. It would be waste of space to point out in detail all slips of this sort. They are, fortunately, irrelevant to the course of the author's argument. But the reader should be warned that these old figures would not pass muster with modern anatomists. -- TRANSLATOR.
[2] This Figure [Fig. 65] would, perhaps, be more nearly typical if the postcentral fissure were separated off from the rest of the fissural complex with which the author has connected it. Again, it is not probable that the calcarine fissure ever appears thus plainly on the lateral surface of the brain (cf. the author's remarks in the text below); though, indeed, but little is certainly known of the fissuration of the occipital lobe. -- TRANSLATOR.