Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3173
(Return to index)
Principles of Physiological Psychology
By Wilhelm Wundt (1902)
Translated by Edward Bradford Titchener (1904)
CHAPTER III
Physiological Mechanics of Nerve-Substance
§1. General Principles and Problems of a Mechanics of Innervation
(a) -- Methods of a Mechanics of Innervation
THE processes that run their course within the elements of the nervous system, the nerve-cells and nerve-fibres described above, have been studied in two different ways. By the one of these, investigators have sought to gain a knowledge of the internal by the other of the external molecular mechanics of nervous substance. The former sets out from an examination of the physical and chemical properties of the nervous elements, and inquires into the changes which these properties evince as a result of physiological function, attempting in this manner to discover the internal forces at work in the nerves and nerve-centres. Inviting as this path may appear, in its promise directly to reveal the intimate nature of the nervous functions, it still takes us so short a distance towards its goal that we cannot venture to trust ourselves upon it. Apart from the scanty results of morphological investigation, mentioned above (p. 53), the study of the functional changes of the central elements is, as yet, hardly more than a programme. And our knowledge of the internal processes in the peripheral nerves is also severely limited. We know that their functioning is attended by electrical and chemical changes, -- the meaning of which is still obscure: we know little more. The only road that remains open to us, therefore, is the second, that of an external molecular mechanics. In taking this, we avoid altogether the question of the special nature of the nervous forces: we set out simply from the proposition that the processes in the elementary divisions of the nervous system are movement-processes, of some sort or other, and that their relations to one another and to the forces of external nature are determined by the mechanical principles valid for motion at large. We thus take up a position akin, let us say, to that of the general theory of heat in modern physics, where the investigator is satisfied to begin with the proposition that heat is a mode of motion, from which -- with the aid of the laws of mechanics -- he derives all the phenomena with systematic completeness. If the molecular mechanics of the nervous system [p. 58] is to accomplish a like result, it must first of all reduce the phenomena that form the subject-matter of its inquiries to their lowest terms: it must investigate the physiological function of the nervous elements, first, under the simplest possible conditions, and, secondly, under conditions that can be experimentally varied and controlled. Now any outside affection of the nervous elements, that serves in some way to arouse or modify their functions, is termed in physiology a stimulus. In using this term, we must, of course, abstract entirely from the ideas which HALLER'S theory of irritability and other modes of thought current in the older vitalistic physiology read into it. If we do this, the term retains its usefulness not only in our modern physiology of the nervous system and its auxiliary organs, but also by extension of meaning in psychology, seeing that all the multiplicity of outside affections that are embraced by it depend primarily upon a peculiar character of living substance itself, and may therefore produce identical results.
Stimuli are classified, in terms of the source from which their activity proceeds, as internal and external. Under internal stimuli are included all stimulatory influences that have their seat in the tissues and organs surrounding the nervous elements: we may instance, especially, rapid changes in the quality of the blood and of the fluids of the tissues. Under external stimuli are included, on the other hand, all the physical and chemical influences exerted upon the organism by the external world in which it lives. As regards nerve-substance, therefore, all stimuli whatsoever are to be classed as external. Whether, for instance, a chemical stimulus arises primarily in the blood in which the nerve-elements are bathed, or makes its way to them from the environment, is indifferent for the intrinsic character of the process. When, however, we desire to apply to nervous substance stimuli of a predetermined intensity and duration, we find, as a rule, that the internal stimuli (in the technical sense) are not available, since they are almost entirely beyond the range of experimental control. We accordingly have recourse to external stimuli, and most frequently to electric shocks and currents, which recommend themselves particularly both by the ease with which they destroy the molecular equilibrium of the nerve-elements, and by the extreme accuracy with which their mode of application may be regulated. In attempting an analysis of the processes in the nerve-fibres, we then begin with that peripheral effect of nervous excitation which is most open to investigation, -- the muscle contraction that follows upon stimulation of the motor nerves, -- and make this our measure of the internal processes. Similarly, for an understanding of the changes in the nerve-cells, we employ the simplest process, amenable to external measurement, that is released in the central organ by the stimulation of a centrally directed nerve-fibre, -- the reflex contraction. In neither of these cases, [p. 59] however, does the muscle-contraction afford a direct measure of the processes that run their course in the corresponding nerve-fibres and at their points of central origin, or of the changes induced in these processes by any determinate outside influence; of itself, it can never furnish more than a certain measure of the processes operative in the substance of the muscle which contracts. As a rule, therefore, every change in the irritability of the nervous elements, to which we have applied artificial stimulation, may be expected to produce a change in the phenomena exhibited by the muscle: thus, if the irritability of the motor nerves is diminished, the muscular contraction will be weaker; if enhanced, it will be stronger. But we shall not be justified in arguing, conversely, that every change in contraction implies a corresponding change in nervous excitability. On the contrary, since the contractile substance has its own intrinsic irritability, which it maintains in face of stimulation whether directly applied or transmitted to it by the motor nerves, very different stimuli may possibly act upon the nerve, or upon the central structures connected with it, to release precisely the same processes in the nervous substance itself, and nevertheless, if the irritability of the contractile substance has changed in the meantime, may produce quite different effects in muscle: or conversely, may set up different processes in the nervous substance, while the contractile substance shows the same reaction. We must, therefore, never lose sight of the fact that the muscular contraction furnishes only an indirect measure of the processes of nervous excitation. If we are to argue immediately from the symptoms of altered contractility to the nervous processes, we must be sure that the observations are made under conditions which guarantee a sufficient constancy in the properties of the muscle experimented upon, or at least make such constancy highly probable. For the rest, the properties of the contractile substance itself, and the related phenomena of the course of the muscular contraction, may here be left out of consideration, as their interest is purely physiological. In no case are we concerned with the muscular contraction save as the changes which it undergoes possess a symptomatic importance for the nervous processes with which they are connected.[1]
It is the task of a physiological mechanics of the nervous substance to reduce the phenomena of nerve-stimulation, so far as they can be traced in the related mechanical phenomena evinced by muscular tissue, to the universal laws of mechanics. In essaying this problem, it must at the outset bring its subject-matter into relation with one, especially, of the great laws of mechanics, -- a law which has proved pre-eminently serviceable [p. 60] in explaining the interrelations of various forms of movement-process. This is the law of the conservation of work.
(b) -- The Principle of the Conservation of Work
We understand by work, in the most general meaning of the term, any operation that changes the position of ponderable masses in space. The amount of work done, in a given case, is accordingly measured by the change of position which it can produce in a weight of determinate magnitude. Ponderable bodies can be moved from their place by light, heat, electricity, magnetism. But all these 'natural forces,' as they are called, are simply forms of molecular motion. It follows, then, that the different modes of molecular motion can do work. The heat of steam, e.g., consists in movements for the most part rectilinear, but oftentimes interferential of the steam particles. As soon as the steam does work, -- let us say, by moving the piston of an engine, -- a corresponding quantum of these movements disappears. This result is commonly expressed in the phrase, 'A certain quantity of heat has been transformed into an equivalent quantity of mechanical work.' It would be more accurate to say that a part of the irregular movements of the steam-particles has been used up, in order to set a larger ponderable mass in motion. We have, then, merely the transformation of the one form of motion into the other; and the work done, measured by the product of the moved weight into the distance through which it is moved, is exactly equal to a sum of lesser amounts of work, which could be measured by the products of the weights of a number of steam-particles into the distances traversed by them, and which now, during the performance of the external work, have disappeared. Conversely, when mechanical work disappears and heat arises in its place, by the friction or compression of physical bodies, we have the opposite transformation of mechanical work into its equivalent amount of molecular work. Not that mechanical work (in the ordinary sense of the term) appears in all cases where heat is latent: the heat is, very commonly, employed simply for the transposition of the particles of the heated body itself. It is a familiar fact that all bodies -- gases most of all liquids and solid bodies in less degree -- expand under the influence of heat. Here, again, molecular work disappears. Just as it is used in the steam-engine to move the piston, so it is used in this case to alter the distance that separates the molecules. Work done in this way is termed work of disgregation. It may be transformed back again into molecular work, as the particles return to their original positions. In general then, molecular work may be transformed either into mechanical action or into work of disgregation, and both of these in their turn may be transformed into molecular work. Now the sum of these three forms of work remains unchanged. This is [p. 61] the, principle of the conservation of work: or, if we choose a name which will permit us, in other contexts, to abstract from that mechanical interpretation of natural processes to which we here stand committed, the principle of the conservation of energy.
This principle is applicable not only to heat, the most general and most widely diffused form of motion, but to other forms as well. In every case, it is always just the one term in the chain of the three interchangeable motions, the character of the molecular work, that is changed. Work of disgregation and mechanical work can be done, e.g., by electricity as well as by heat. There are, therefore, various kinds of molecular work; but there is in the last resort only one work of disgregation, as there is only one form of mechanical work. Disgregation is the name given, in every instance, to a permanent change of the distances separating the molecules, no matter what cause has produced it. When we distinguish a simple increase in the volume of a body from a change of its aggregate condition, and this again from chemical decomposition, or dissociation, we are really distinguishing nothing more than three degrees of disgregation. Mechanical work, in the same way, consists always and everywhere in the change of position of ponderable masses. It should be noted that the different forms of molecular work may also, under certain circumstances, be transformed into one another. Thus, a certain quantum of electrical work may give rise, simultaneously, to heat, disgregation and mechanical work.
It is from mechanical work that the idea of work, in the abstract, has been derived. And it is mechanical work that is selected, from the various forms of work mentioned above, to serve as a common measure of work at large. The reason is, that mechanical work can be most accurately measured, and that the only possibility of a comparison of the different forms of work is given with the reduction of all to one. This measure, now, is applied in the special case by help of the principle of the conservation of work, which lays it down that a given amount of molecular work or work of disgregation is equivalent to the mechanical work into which it is transformed or from which it is generated. In the performance of mechanical work, a ponderable body may be lifted, against the force of gravity, or moved by its own weight, or accelerated in spite of friction, and so on. In the latter event, the portion of mechanical work necessary to overcome friction is transformed into heat. Where the body is lifted, we suppose that the work employed for the lifting is stored up within it, since this work call be passed on again to other bodies, by a subsequent fall of the weight from the same height. Disgregation behaves, in this regard, just as the lifted weight does: a certain quantity of molecular work, mostly in the form of heat, is used up in its production, and this same quantity must reappear as soon as the disgregation is abolished. But a lifted weight remains lifted [p. 62] so long as its weight is held in equilibrium by some other form of work, e.g. by the heat-motion of expanded steam. In the same way, the disgregation of the molecules of a body persists, so long as their reunion is prevented by some form of internal work, e.g. by heat-vibrations. Hence, between the moment at which the weight is lifted or the disgregation of the molecules effected, and the moment at which the work required for these operations is reproduced by the fall of the weight or the union of the molecules, there may intervene a static condition, continuing for a longer or shorter time, throughout which just so much internal work is being done as is necessary for the maintenance of equilibrium, -- so that no alteration takes place in the existing status, in the position of bodies and their molecules, in temperature, in electrical distribution. Only at the moment when this state of equilibrium is disturbed, when the weight falls or the molecules approach one another, do transformations of work set in again. The mechanical work or work of disgregation is now transformed first of all into molecular work, usually into heat, and this may in its turn pass over in part into mechanical action or disgregation of molecules; the transformations continuing, until circumstances occur that favour the reinstatement of the stationary condition. Since, now, there is a certain sum of work available, in a lifted weight or in disgregated molecules, we may consider every lifted weight and every disgregation as potential work or work of position. The amount of this potential work is always precisely the same as the amount of work that was required to effect the lift or the disgregation, and as the amount of work that may reappear in consequence of fall or of aggregation. The law of the conservation of work may, accordingly, be expressed in other terms as follows: the sum of actual and of potential work, of work of position and work of motion, remains constant. It is clear that this is only a special way of formulating our previous law of the conservation of the sum of work; for we always mean by work of position a lift or a disgregation accomplished by expenditure of actual work, and maintained by a stationary condition of tension or motion. If we could observe the smallest oscillatory movements of the atoms as well as the motions of bodies and the permanent changes of position that they undergo, the law would hold of these atomic movements also, that the sun of actual and potential work remains unchanged. In actual fact, however, where the particles of the mass are in constant motion about approximately the same positions of equilibrium, matter appears to us to be at rest. We accordingly term the work done, invisibly to us, in a stationary condition, 'internal' molecular work, thus distinguishing it from the molecular work which arises when there is a change in the state of equilibrium as regards temperature, electric distribution, etc., and which we call 'external' molecular work.[p. 63]
These stationary conditions are continually alternating with changes of state. The stage of nature is thus occupied, in never-ending succession, with the passage from internal to external and from external back again to internal molecular work. It will suffice here to give illustrations of the processes that have the most direct bearing upon our own problem, -- illustrations of disgregation and its reversal. Differences in aggregate condition depend, it is supposed, upon different states of molecular motion. The molecules of a gas repel one another, and consequently continue to move, in rectilinear paths, until such time as they strike the wall of the containing vessel, or other molecules, from which they rebound. In liquids, the molecules oscillate about instable, in solids, about stable positions of equilibrium. If, now, we are e.g. to transform a liquid into a gas, we must increase the work of the molecules. We do this by the application of heat. So long as only the molecular work of the liquid increases, nothing results but an increase of its temperature. But if, at the same time, we allow the liquid to expand, then a part of its molecular work is further transformed into disgregation. Finally, if the application of heat is continued, and the disgregation carried to the point at which the particles of the liquid travel beyond the spheres of their mutual attraction, the liquid is suddenly transformed into gas or steam: it now enters upon a new state of equilibrium, in the production of which a large amount of molecular work, i.e. of heat, has been consumed. If heat is now withdrawn from the steam, so that its internal work is diminished, a point will be reached, on the backward path, at which the average distance between the molecules is sufficiently reduced to bring them once more within the limits of their mutual attraction. With the supervention of this original position or equilibrium, molecular work must be done, i.e. heat be liberated, as a result of the renewed activity of the forces of attraction; and the amount of heat thus disengaged is precisely the same as that consumed in the first instance.
What holds in this case holds, in practically the same way, for the decomposition and recomposition of chemical compounds. In every substance we can distinguish between the state of physical and the state of chemical equilibrium. For every molecule, in the physical sense, consists of a number of chemical molecules or (to use the term applied to the indecomposable chemical molecule) of a number of atoms. Just, then, as the molecules may exist in different conditions of motion, varying with the aggregate state of the body in question, so may the atoms also, according to the character o the chemical compound. Modern chemistry regards all bodies as compounds; chemically simple bodies are looked upon as compounds of homogeneous atoms. Hydrogen gas is thus every whit as much a chemical compound as is hydrochloric acid: in the former, [p. 64] two atoms of hydrogen are compounded together (H.H), in the latter, one atom of hydrogen is compounded with one of chlorine (H.Cl). Here again, however, what appears to be matter at rest is in reality only a stationary condition of motion. The atoms in a chemical compound oscillate, it is supposed, about more or less stable positions of equilibrium. The character of this motion is, at the same time, strongly influenced by the aggregate condition of the compound, regarded as a physical body. Thus, in gases and liquids, the state of motion of the chemical atoms is, as a rule, comparatively free; atoms are occasionally torn from their connexions, and at once compound again with other atoms that have been similarly released. In hydrochloric acid, for instance, gaseous or liquid, the average composition of all chemical molecules is HCl. Nevertheless, separate atoms H and Cl are constantly occurring in the free state, though they cannot maintain it, but are always compelled at once, by the forces of chemical attraction, to enter again into combination. From this point of view we gain a satisfactory explanation of the ready decomposability of gases and liquids in face of heat, electricity and other chemical compounds.[2]
We find, once more, in the aggregation of chemical molecules, differences analogous to those which we have noted in the aggregate states of physical bodies. There are relatively stable and relatively instable chemical compounds. In the former, the forces of attraction, in virtue of which the particles vibrate about certain determinate positions of equilibrium, are stronger; in the latter, weaker. These differences of chemical aggregation are, of course, altogether independent of the physical since the physical molecules are always, to start with, chemical aggregates. Very stable compounds may accordingly occur in the gaseous state, and very unstable in the aggregate state of solidity. In general the compounds of homogeneous atoms, the chemically simple substances, belong to the less stable compounds; most of them, certain of the metals excepted, decompose fairly easily to form compounds with heterogeneous atoms. The same thing is true, on the other hand, of extremely complex compounds, which readily break up into simpler. Here belong most of the 'organic' substances. It follows, then, that stable chemical compounds are to be found predominantly among the simpler connexions of heterogeneous atoms. Thus, carbonic acid, water, ammonia, and many of the metallic oxides and inorganic acids are decomposed only with difficulty. Just, however, as the different aggregate states can be transformed into one another, so may relatively instable compounds be transformed into stable, and conversely. There is, as ST. CLAIRE DEVILLE proved, no compound so stable that it cannot be dissociated by the application of heat in sufficient quantity. Here, as in the change of a liquid into a gas, a certain amount of the [p. 65] internal work of the heat disappears, transformed into work of dissociation. When the dissociation is complete, the atoms are in a new state of equilibrium. In the dissociation of water, e.g., the more stable connection H2O gives place to the less stable forms H.H and O.O, in which the vibratory condition of the atoms differs from that in the stable compound H2O very much as the vibratory condition of the molecules of steam differs from that of the molecules of water; that is to say, the atoms in their new, instable connexions will, on the whole, describe longer paths, and consequently do more internal molecular work. To make up this deficiency, heat is necessary. The work thus expended upon dissociation is, however, still present as potential work: for when the new state of equilibrium of the dissociated molecules is disturbed, they are able to compound again, and the work of dissociation once more makes itself apparent in the form of heat. The chemical molecules have, at the same time, passed into their former condition of equilibrium, where the stationary work which they perform in movements about their positions of equilibrium is diminished by the amount of the internal work released in the act of composition. We see, then, that the phenomena connected with composition and dissociation are identical with the phenomena observed in the alternation of aggregate states, save only that much larger amounts of work are usually required for dissociation than for disgregation, and that in the former case the exchange between work of position and work of motion attains proportionately higher values.
(c) -- Application of the Principle of the Conservation of Work to the Vital Processes of the Nervous System
The tissues of the living organism are the seat of chemical processes which, by their great regularity of occurrence, furnish a remarkable illustration of the alternations of potential and actual internal and external work. In the plants, we have a constant dissociation of stable compounds. Carbonic acid, water, ammonia, the nitric acid and sulphuric acid of the nitrates and sulphates, are taken up by the plant, and decomposed into less stable compounds -- wood fibre, starch, sugar, albumins, etc. -- in which a large amount of potential work is stored; at the same time, oxygen is eliminated. These compounds, produced by the plant, are transformed in the animal body, by help of atmospheric oxygen (i.e. by a process of combustion), into the more stable compounds from which the plant had derived them; at the same time, the potential work stored up in the organic compounds goes over into actual work, partly in the form of heat, partly in that of external work of the contractile substance. The central station, from which all these processes of the animal body are directed, is the nervous system. [p. 66] It maintains the functions that subserve the processes of combustion; it regulates the distribution and radiation of heat; it determines the activity of the muscles. In many cases, it is true, and especially in cases of muscular action, the issuance of impulses from the nervous system is itself directed by external movement-processes, the sense-stimuli. The true source of its functional capacity lies, however, not in these, but in the chemical compounds of which nerve-mass and contractile substance are composed, and which are taken over, almost without modification, from the living laboratory of plant-tissue. These contain the store of potential work, which under the influence of external stimulation is transformed into actual work.
The compounds of which the nerve-mass consists remain, so long as stimulus-processes do not intervene to modify them, approximately in that stationary condition which appears to outward observation as a state of rest. This rest is, however, here, as in all such instances of a stationary condition, only apparent. The atoms of the complex chemical compounds are in continual motion; now and again, they travel beyond the sphere of operations of the atoms with which they have hitherto been combined, and come within that of other atoms, freed like themselves. There is, therefore, in a liquid so easily decomposable as the nerve-mass, a constant alternation of decomposition and recomposition of chemical compounds, and the mass appears stationary simply for the reason that, on the average, there are as many processes of the one kind going on as of the other. In this particular instance, however, we cannot in strictness say even so much: not even during their period of rest is the state of the nervous elements really constant and unchanged. With compounds of such complexity, it invariably happens that certain of the atoms which have been removed from their former sphere of operations do not, in reuniting, enter into their old connexions, or into connexions of the same order, but combine afresh to form similar and more stable compound. This process is termed intrinsic decomposition. In the living organism the disturbances arising from intrinsic decomposition are compensated by the removal of the products of decomposition, and by the intake of new materials for the renewal of the constituents of the tissues.
We may, then, consider resting nervous substance as a semisolid mass given in a stationary condition of motion. In such a mass, there is no release of external work; the work values produced by the individual atoms cancel one another. This cancellation takes place, in large measure, within the complex chemical molecules. As the atoms of the molecule oscillate about their positions of equilibrium, each one of them does a certain work, which, however, is counteracted by the work of other atoms, and consequently is not perceptible outside the molecule. This internal [p. 67] molecular work is far more considerable in an instable chemical compound, owing to the greater freedom of movement possessed by the atoms, than it is in a stable compound. It is this, therefore, that represents the potential work of the compound. For if the existing state of equilibrium be disturbed, the relatively instable may pass into a relatively stable compound; in which event the surplus of internal molecular work contained in the former is at once transformed into external. To a certain extent, however, the establishment of equilibrium takes place without the chemical molecule. Where atoms are continually passing from less stable to more stable connexions, work must appear: where, on the other hand, atoms are transferred from more stable to less stable connexions, work must correspondingly disappear: and in both cases it is external molecular work, generally heat, that is produced and consumed again. We may term the work that appears with the origination of the more stable compound 'positive' molecular work, and the work that disappears with the formation of the less stable compound, 'negative' molecular work. The condition of true equilibrium in a decomposable liquid like the nerve-mass will then be this: that the internal molecular work or potential work be kept unchanged, by the continual compensation of the existing quantities of positive and negative external molecular work. Or, to but the same thing in different words: the internal molecular work must be kept constant by the renewal (through retransformation into internal molecular work) of all that it loses in external molecular work. What changes, now, are brought about in this stationary condition of the nerve by the development of the process of stimulation?
§2. The Course of the Processes of Stimulation in the Nerve-Fibre
(a) -- Course of the Muscular Contraction following Stimulation of the Motor Nerve
The simplest of all the external phenomena that can inform us of the nature of the processes of stimulation in nerve is the muscular contraction which sets in, and runs its course in time, as a result of stimulation of a motor nerve. Fig. 24 shows the 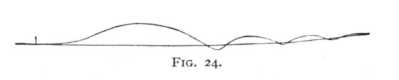 course of a contraction of this kind in the gastrocnemius of the frog; a lever with writing-point was attached to the muscle, and recorded the phases of the contraction directly upon a quickly moving smoked-glass plate, carried by a heavy pendulum. The conditions under which the tracing was obtained were made as simple as possible, in order that the course of the contraction might really be symp-[p. 68]tomatic of the stimulatory process. The muscle carried no weight beyond the light writing lever, and gave a twitch in response to stimulation. Numerous observations have shown that the loading of a muscle increases its irritability. Under the present circumstances this intensifying effect may be regarded as relatively small and the influence that it exerts upon the various experiments whose results we are to compare, as sufficiently uniform.[3] The vertical stroke to the left indicates the moment at which the stimulus was applied to the nerve. The resulting curve, whose axis of abscissas appears by reason of the movement of the pendulum as an are of a circle, shows that the twitch sets in perceptibly later than the stimulation, and that the contraction rises at first quickly, then more slowly, to be followed in like manner by a gradual relaxation. If the stimulus is momentary, the whole twitch is generally completed in 0.08-0.1 sec. Provided that the nerve is stimulated directly above the muscle, about 0.01 sec. of this time is lost between the application of stimulus and the beginning of the twitch; this interval is known as the stage of latent stimulation, or the latent period. The experiment makes it probable that the movement process in nerve is relatively slow. Since, however, we have not determined how much of this retardation of the processes is referable to the inertia of the muscular substance, the result obtained is not of decisive value.
course of a contraction of this kind in the gastrocnemius of the frog; a lever with writing-point was attached to the muscle, and recorded the phases of the contraction directly upon a quickly moving smoked-glass plate, carried by a heavy pendulum. The conditions under which the tracing was obtained were made as simple as possible, in order that the course of the contraction might really be symp-[p. 68]tomatic of the stimulatory process. The muscle carried no weight beyond the light writing lever, and gave a twitch in response to stimulation. Numerous observations have shown that the loading of a muscle increases its irritability. Under the present circumstances this intensifying effect may be regarded as relatively small and the influence that it exerts upon the various experiments whose results we are to compare, as sufficiently uniform.[3] The vertical stroke to the left indicates the moment at which the stimulus was applied to the nerve. The resulting curve, whose axis of abscissas appears by reason of the movement of the pendulum as an are of a circle, shows that the twitch sets in perceptibly later than the stimulation, and that the contraction rises at first quickly, then more slowly, to be followed in like manner by a gradual relaxation. If the stimulus is momentary, the whole twitch is generally completed in 0.08-0.1 sec. Provided that the nerve is stimulated directly above the muscle, about 0.01 sec. of this time is lost between the application of stimulus and the beginning of the twitch; this interval is known as the stage of latent stimulation, or the latent period. The experiment makes it probable that the movement process in nerve is relatively slow. Since, however, we have not determined how much of this retardation of the processes is referable to the inertia of the muscular substance, the result obtained is not of decisive value.
We come closer to the movement in nervous substance itself when we stimulate the nerve at two different points of its course, the one remote from the muscle, the other as near to it as possible, and when the experiment is so arranged that the stimulation 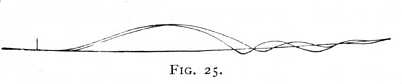 is timed to occur in both cases at the same point upon the axis of abscissas above which the curve of contraction is described. If the two stimuli have the same intensity, and the nerve is kept in as constant a condition as possible, the resulting curves evince a twofold difference. In the first place, as HELMHOLTZ discovered, the curve of contraction given by the more remote stimulus begins later -- has [p. 69] a longer latent period -- than the other. Secondly, as PFLÜGER first showed, the twitch released higher up the nerve is the stronger; its curve is higher and also, as the author pointed out, of longer duration. If, therefore, the experimenter desires to obtain two muscle-curves of the same height, he must apply a somewhat weaker stimulus to the part of the nerve that is more remote from the muscle. Even then it usually happens, provided the experiment be made on the living animal, that the corresponding contraction lasts for a little longer time. The two curves will accordingly differ in the manner indicated in Fig. 25. There is a brief interval between the starting-points of the contractions, which evidently corresponds to the time which the excitation requires for propagation from the upper to the lower point of stimulation; and the twitch released higher up, although in this case it was excited by a weaker stimulus, reaches the axis of abscissas later than its initial retardation would lead us to expect. We may, then, conclude from these experiments, first, that the movement-process of stimulation is relatively slow, -- for the frog-nerve at ordinary summer temperature it averages 26, for the nerves of warm-blooded animals at normal body temperature 32 m. in the 1 sec., -- and, secondly, that it consists, in all probability, not in a simple transmission and propagation of the external stimulus movement, but in a chain of movement-processes released from one point to another within the nerve itself. This latter inference is borne out, more particularly, by the lengthening of the contractions which goes with increased distance of the point of stimulation from the muscle. The phenomenon is altogether constant, and may be observed most strikingly in the uncut nerves of the living animal.[4]
is timed to occur in both cases at the same point upon the axis of abscissas above which the curve of contraction is described. If the two stimuli have the same intensity, and the nerve is kept in as constant a condition as possible, the resulting curves evince a twofold difference. In the first place, as HELMHOLTZ discovered, the curve of contraction given by the more remote stimulus begins later -- has [p. 69] a longer latent period -- than the other. Secondly, as PFLÜGER first showed, the twitch released higher up the nerve is the stronger; its curve is higher and also, as the author pointed out, of longer duration. If, therefore, the experimenter desires to obtain two muscle-curves of the same height, he must apply a somewhat weaker stimulus to the part of the nerve that is more remote from the muscle. Even then it usually happens, provided the experiment be made on the living animal, that the corresponding contraction lasts for a little longer time. The two curves will accordingly differ in the manner indicated in Fig. 25. There is a brief interval between the starting-points of the contractions, which evidently corresponds to the time which the excitation requires for propagation from the upper to the lower point of stimulation; and the twitch released higher up, although in this case it was excited by a weaker stimulus, reaches the axis of abscissas later than its initial retardation would lead us to expect. We may, then, conclude from these experiments, first, that the movement-process of stimulation is relatively slow, -- for the frog-nerve at ordinary summer temperature it averages 26, for the nerves of warm-blooded animals at normal body temperature 32 m. in the 1 sec., -- and, secondly, that it consists, in all probability, not in a simple transmission and propagation of the external stimulus movement, but in a chain of movement-processes released from one point to another within the nerve itself. This latter inference is borne out, more particularly, by the lengthening of the contractions which goes with increased distance of the point of stimulation from the muscle. The phenomenon is altogether constant, and may be observed most strikingly in the uncut nerves of the living animal.[4]
In order, now, to gain a deeper insight into the course of the phenomena of stimulation, we must endeavour to inform ourselves of the state of the nerve at each successive moment of the time following upon stimulation. We may do this, always in terms of the external effects of nervous activity, by investigating the behaviour of the nerve, at every moment of the period of stimulation, in face of a second, test-stimulus of constant magnitude.[p. 70] Here, in the case of the simple muscle-contraction, the properties of the muscular substance itself naturally contribute their share to the total result. We can, however, eliminate their influence, very much in the same way that we did in the experiments on the propagation of stimulation. Where the conditions residing in the muscle remain contact, the observed changes must necessarily depend upon the processes taking place in the nerve.
(b) -- Excitatory and Inhibitory Processes in Nerve-Stimulation
We must suppose, if we apply the principle of conservation to the processes in the nerve, that every processes of stimulation produces two opposite effects in the nerve-fibre. The one set of operations will be directed upon the production of external work (muscular contraction, development of heat, secretion, stimulation of nerve-cells), and the other upon the recovery of the work thus liberated. We may term the former the excitatory, and the latter the inhibitory effects of stimulation. The whole course of the stimulation is then dependent upon the constantly varying play of excitation and inhibition. In order to demonstrate, by means of our test-stimulus, which of these processes, excitation or inhibition, has the upper hand, we may employ either of two different methods. We may work with stimulation-processes of so little intensity that they are unable of themselves, without the intervention of the test-stimulus, to release any muscular contraction at all; or we may eliminate the influence of the contraction itself during the time of its occurrence. We can do this, in cases where we are concerned to demonstrate an increase of irritability, by overloading the muscle, i.e. by attaching to it so heavy a weight that both the original twitch and the contraction normally released by the test-stimulus are suppressed, or at most only a minimal (what is called an 'isometric') twitch remains possible. If now, during the process of the first stimulation, the test-stimulus nevertheless releases a more than minimal contraction, we have evidence of an increase of the excitatory effects and, in the height of the muscle-curve, a rough measure of their magnitude. gives an illustration of this procedure. The stimulation-process here under 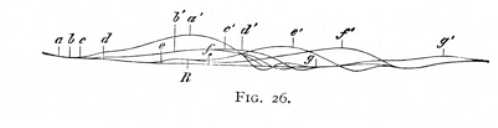 investigation was set up by the closing of a constant current in the ascending direction: the positive electrode, that is, lay nearer to the muscle, and the negative farther away from it. The current was closed at the point a. In response to the stimulation, the muscle (not overloaded) gave the twitch [p. 71] recorded as a'. The load was now attached, and the muscle-curve reduced by it to the minimal height R. The test-stimulus, employed to test the state of the nerve in successive phases of the stimulation-process, was the break shock of an induction-current, applied a short distance below the length of nerve stimulated by the constant current. So long as the latter was open, the twitch produced by the shock in the overloaded muscle was also minimal. A series of experiments was then preformed, in which the nerve of the overloaded muscle was first of all stimulated at a by make of the constant current, and then again, after a definite period, by application of the test-stimulus. If the two stimuli were coincident (a), the height of the muscle-curve remained minimal. Where the test-stimulus came later, the successive times of stimulation b, c, d, e, f, g, gave the contractions b', c', d', e', f', g'. The course of these curves shows clearly that the stimulated nerve undergoes a change of state, which manifests itself as an increased irritability. The change begins shortly after the stimulation a; reaches a maximum that corresponds approximately with the highest points of the contractions a' and R (e e'); and then gradually decreases again, -- though it persists, as is shown by the final test g g', for a considerably longer time than the primary twitch a'.
investigation was set up by the closing of a constant current in the ascending direction: the positive electrode, that is, lay nearer to the muscle, and the negative farther away from it. The current was closed at the point a. In response to the stimulation, the muscle (not overloaded) gave the twitch [p. 71] recorded as a'. The load was now attached, and the muscle-curve reduced by it to the minimal height R. The test-stimulus, employed to test the state of the nerve in successive phases of the stimulation-process, was the break shock of an induction-current, applied a short distance below the length of nerve stimulated by the constant current. So long as the latter was open, the twitch produced by the shock in the overloaded muscle was also minimal. A series of experiments was then preformed, in which the nerve of the overloaded muscle was first of all stimulated at a by make of the constant current, and then again, after a definite period, by application of the test-stimulus. If the two stimuli were coincident (a), the height of the muscle-curve remained minimal. Where the test-stimulus came later, the successive times of stimulation b, c, d, e, f, g, gave the contractions b', c', d', e', f', g'. The course of these curves shows clearly that the stimulated nerve undergoes a change of state, which manifests itself as an increased irritability. The change begins shortly after the stimulation a; reaches a maximum that corresponds approximately with the highest points of the contractions a' and R (e e'); and then gradually decreases again, -- though it persists, as is shown by the final test g g', for a considerably longer time than the primary twitch a'.
Where it is not, as in the instance here taken, the excitatory, but the inhibitory effects that have the upper hand, the method of overloading naturally ceases to be applicable. We can, however, readily infer the presence of inhibitory influences from the magnitude of the effect produced by the test-stimulus during the progress of the contraction. If, e.g., the test-stimulus produces no effect whatever, we can argue with perfect certainty to the preponderance of inhibitions. An illustration of this state of things is given in Fig. 27. The stimulation process here under investigation was again set up by the making of an ascending constant current; and the test-stimulus was, as before, the break shock of an induction-current applied below the portion of nerve stimulated by the constant current. A and B represent two successive experiments, in each of which the current was closed at a and the test-stimulus thrown in at b. The primary object of both experiments was to investigate, first, the effect of the current without the test-stimulus, and secondly, the effect of the test-stimulus without preceding closure of the constant current: this 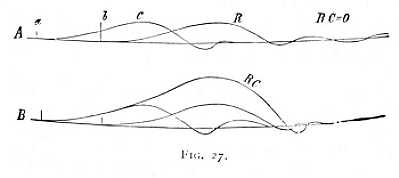 gave the contractions C and R, which are precisely alike in A and B. In the next place, the test-stimulus was applied, at b, immediately after the closure of the constant current at a. The results obtained in the experiments A and B were now entirely different. In A, a simple contraction C was recorded, precisely as if the test-stimulus R had not operated at all (RC=0); in B, the curve of contraction was at first coincident with C, but, when the time came for the beginning of the contraction R, rose so far above C that RC is higher than the curves R and C [p. 72] taken together. From this difference of result we may conclude that in A a strong inhibition persisted during the progress of the stimulation C, while in B there was either a preponderance of excitatory effects or no change of irritability at all. To decide between these alternatives, we have only to overload the muscle, in the manner indicated above, and so to reduce the contractions C and R to zero or to minimal height. Adopting this method, we find that, as a matter of fact, in experiment R the excitatory effects had the upper hand. -- Now the difference between the experimental conditions of A and B was this: that in A the test-stimulus was applied very near the part of the nerve stimulated by the constant current, while in B it lay nearer the muscle. Hence the experiments show that, in one and the same process of stimulation, the inhibitory effects may predominate in one portion of a nerve, and the excitatory in another.[5]
gave the contractions C and R, which are precisely alike in A and B. In the next place, the test-stimulus was applied, at b, immediately after the closure of the constant current at a. The results obtained in the experiments A and B were now entirely different. In A, a simple contraction C was recorded, precisely as if the test-stimulus R had not operated at all (RC=0); in B, the curve of contraction was at first coincident with C, but, when the time came for the beginning of the contraction R, rose so far above C that RC is higher than the curves R and C [p. 72] taken together. From this difference of result we may conclude that in A a strong inhibition persisted during the progress of the stimulation C, while in B there was either a preponderance of excitatory effects or no change of irritability at all. To decide between these alternatives, we have only to overload the muscle, in the manner indicated above, and so to reduce the contractions C and R to zero or to minimal height. Adopting this method, we find that, as a matter of fact, in experiment R the excitatory effects had the upper hand. -- Now the difference between the experimental conditions of A and B was this: that in A the test-stimulus was applied very near the part of the nerve stimulated by the constant current, while in B it lay nearer the muscle. Hence the experiments show that, in one and the same process of stimulation, the inhibitory effects may predominate in one portion of a nerve, and the excitatory in another.[5]
We must not omit to mention the fact that, in all these cases, it depends upon the nature of the test employed whether the one or the other of the opposed effects, the excitatory or the inhibitory, is the more clearly demonstrable. Weak stimuli are, without exception, better for the proof of inhibition, strong stimuli for that of excitation. If, however, we test the same stimulation-process with weak and strong stimuli alternately, we find in most instances that, during the greater part of its course, the excitatory and the inhibitory effects are both alike enhanced. At a phase of the stimulation-process when the effect of weak test-stimuli is wholly suppressed, the effect of strong stimuli may be increased.[6]
It follows from the above results that, if we desire to gain a quantitative expression of the relation which the inhibitory effects hear at any given moment to the excitatory, we shall best have recourse to 'isometric' contractions and to stimuli of moderate intensity, that are, on the whole,[p. 73] equally sensitive to inhibition and to excitation. Experiments made under these conditions show that tee stimulation-process developing as the result of a momentary stimulus, e.g. of an electric shock or mechanical concussion, runs its course as follows. At the moment of stimulation, and for a brief period afterward, the nerve does not react to the weak test-stimulation at all; the process takes precisely the same form as it would if the stimulus had not acted.[7] If, therefore, we apply to the same point upon the nerve or to two neighbouring points, first a stimulus R (Fig. 28), then a stimulus C, and finally the two stimuli R and C together, the curve RC recorded in the third case is identical with the more intensive of the two single-stimulus curves R, C: in our illustration, with R (Fig. 28 A). We obtain the same result if we allow a very brief interval to elapse between the times of stimulation a, b. So soon as this interval becomes noticeable, however, the combined stimuli provoke a stronger contraction than either of them gives separately. Even while the time difference is less than the ordinary latent period, it not uncommonly happens that RC is greater than the sum of R and C: taken together; and the more nearly minimal the contractions, the greater does the excess become (Fig. 28 B). This enhancement of irritability increases up 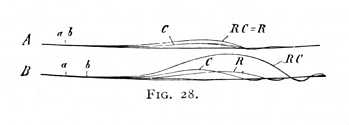 to a point corresponding roughly with the maximum of contraction, and then gives place to a decrease; at the same time, it can be demonstrated for a considerable period after the conclusion of the twitch. Fig. 26 (p. 70 above) gives a picture of the whole process. We may say, then, in summary, that the course of the stimulation-process is in general divisible into three stages: the stage of inexcitability: the stage of increasing, and the final stage of decreasing excitability.
to a point corresponding roughly with the maximum of contraction, and then gives place to a decrease; at the same time, it can be demonstrated for a considerable period after the conclusion of the twitch. Fig. 26 (p. 70 above) gives a picture of the whole process. We may say, then, in summary, that the course of the stimulation-process is in general divisible into three stages: the stage of inexcitability: the stage of increasing, and the final stage of decreasing excitability.
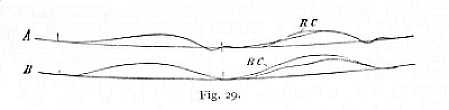
Oftentimes, however, this third stage is interrupted by a brief interval, during which irritability shows a sudden marked decrease, quickly followed by another increase. The decrease always coincides with the conclusion of the twitch. It passes so rapidly, that it can be recognised only by the increase of the latent period of the test-stimulus; and it is of regular occurrence only where the functional capacity of nerve and muscle is very high. An instance of this transitory inhibition at the conclusion of the twitch is given in Fig. 29 A. The contraction to the left corresponds to the stimulation process under investigation; the unlettered twitch to the right is the result of simple application of the test-stimulus; while RC is the twitch released by the test-stimulus under the influence of previous stimulation.[p. 74] The curves of A were obtained from a fresh nerve, those of B from a nerve that had already been subjected to repeated stimulation.[8] In this phenomenon, the period following the conclusion of the twitch forms a precise parallel to the latent period preceding contraction. In both these cases, however, it is not impossible that the result is in
 some measure due to conditions residing in the muscle itself. Thus, the stage of inexcitability, which appears at the beginning of stimulation, may very possibly be attributable to the fact that the contractile substance requires a certain time to initiate a contraction. In the same way, the stage of diminished excitability that coincides with the conclusion of the twitch may be explained by the assumption that opposing influences within the muscle -- already at work, perhaps, during the rapid progress of the contraction -- now operate in full force. Nevertheless, the reactive effect in both stages alike must, in all probability, be regarded as a phenomenon for which nerve and muscle are jointly responsible. This view is borne out by the fact that the duration of the two stages of inhibition is largely determined by the character of the stimuli which affect the nerve. If, e.g., we apply a stimulus to a portion of nerve that lies within the sphere of operation of the anode of a constant current, the duration of the inhibitory stages is considerably lengthened.
some measure due to conditions residing in the muscle itself. Thus, the stage of inexcitability, which appears at the beginning of stimulation, may very possibly be attributable to the fact that the contractile substance requires a certain time to initiate a contraction. In the same way, the stage of diminished excitability that coincides with the conclusion of the twitch may be explained by the assumption that opposing influences within the muscle -- already at work, perhaps, during the rapid progress of the contraction -- now operate in full force. Nevertheless, the reactive effect in both stages alike must, in all probability, be regarded as a phenomenon for which nerve and muscle are jointly responsible. This view is borne out by the fact that the duration of the two stages of inhibition is largely determined by the character of the stimuli which affect the nerve. If, e.g., we apply a stimulus to a portion of nerve that lies within the sphere of operation of the anode of a constant current, the duration of the inhibitory stages is considerably lengthened.
We are now in a position to discuss the relation of the excitatory and inhibitory effects within the nerve fibre, in abstraction from the properties accruing to the reacting muscle. We may conceive of them as follows. On the occurrence of stimulation, excitatory and inhibitory effects are produced simultaneously. At first, the latter are very much the stronger. As time goes on, however, they increase more slowly, while the excitatory effects advance more quickly. Oftentimes, these last appear to maintain their ascendancy until the conclusion of the whole process. If however, the functional capacity of the: nerve is very high, the inhibitory effects may again acquire the upper hand for a brief period immediately following the conclusion of the twitch. This fact indicates, at the same time, that the process is not entirely continuous, but that the rapid result produced in contraction by the excitatory effects is always followed by an inhibitory reaction. The release of excitation thus resembles a sudden discharge, in [p. 75] which the available forces are quickly consumed, so that for a short time the opposite effects are in the preponderance. Fig. 30 is an attempt to show this sequence of events in graphic form. Stimulation occurs at rr'. The curve ab represents the course of the excitatory, the curve cd of the inhibitory effects, measured by the magnitude of the downward directed (negative) ordinates of the curve cd. We assume that excitatory and inhibitory impulses are already present in 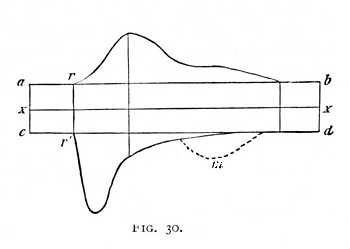 the nerve, before the application of stimulus, but that they are in equilibrium. These pre-existent impulses we make proportional to the ordinates xa and xc. The curve of inhibition is characterised by a rapid rise at its commencement, the curve of excitation by the gradual fall at its conclusion. What we term the functional capacity of a nerve is a function at once of excitation and of inhibition. The more functionally capable the nerve, the greater is the efficacy of both the inhibitory and the excitatory forces contained within it. In the exhausted nerve, both alike are diminished, but the inhibitory in higher degree. Here, therefore, the irritability is enhanced, and the transitory inhibitions at the conclusion of the twitch, which may perhaps be referred (as indicated by the dotted curve m) to an oscillatory repetition of the inhibitory process, are no longer observable.
the nerve, before the application of stimulus, but that they are in equilibrium. These pre-existent impulses we make proportional to the ordinates xa and xc. The curve of inhibition is characterised by a rapid rise at its commencement, the curve of excitation by the gradual fall at its conclusion. What we term the functional capacity of a nerve is a function at once of excitation and of inhibition. The more functionally capable the nerve, the greater is the efficacy of both the inhibitory and the excitatory forces contained within it. In the exhausted nerve, both alike are diminished, but the inhibitory in higher degree. Here, therefore, the irritability is enhanced, and the transitory inhibitions at the conclusion of the twitch, which may perhaps be referred (as indicated by the dotted curve m) to an oscillatory repetition of the inhibitory process, are no longer observable.
(c) -- After-effects of Stimulation: Practice and Fatigue
Our study of the changes of excitability which take place in a nerve during the process of stimulation has shown two things: that the effect produced in the nerve disappears not abruptly but quite gradually, and that it always persists for a noticeable time after the conclusion of the twitch. We have now to consider another phenomenon, which evidently proceeds from the same causes. If several stimuli are successively applied at such intervals that each falls within the period of decline of the stimulation set up by its predecessor, the irritability of the nerve is increased. Indeed, under favourable conditions this increase may be so considerable that a weak stimulus, which at first could not provoke any contraction at all finally releases a maximal contraction. At the same time, the contractions become longer; while the longer after-effect shows further that the course [p. 76] of the excitation has increased not only in intensity but also in duration. These phenomena occur both with stimulation by electric shocks and with instantaneous mechanical stimuli. They are therefore bound up with the stimulation process as such; although, where electrical stimuli are employed, they undergo modification as a result of certain processes developed at the two electrodes: these processes, which we shall discuss presently, are essentially different at anode and cathode.[9] If, on the other hand, we apply the stimuli in very quick succession, so that the twitch provoked in any given case begins before the twitch released by the preceding stimulus has run its full course, we obtain the permanent contraction known by the name of tetanus, and consisting essentially of a summation of the super-posed twitches.[10] This summation of contractions is here of no further interest to us; we note simply that the properties of the contractile substance have an important part to play in its origination. If we abstract from it, we may say that the phenomena of increased excitability in consequence of preceding stimuli, which we are now considering, are in the main indicative of the behaviour of the nervous substance; the muscle is, in all probability, concerned in their production only in so far as it resembles nerve in the general character of its irritability. This conclusion is borne out more especially by the fact that the increase of excitability by stimulation is independent of the occurrence of contraction. Where the stimulus employed is so weak that it cannot release any contraction at all or where the muscle is so overloaded that the contraction is entirely suppressed, the increase of excitability is just as noticeable as it is when the muscle is allowed to contract. Nay more: since, under these circumstances, the phenomena of fatigue (which we discuss below) are ruled out, it becomes as a general rule still more noticeable. Taking the whole group of facts into account, we may therefore designate this increase of excitability as the elementary phenomenon of the process of practice. For when we speak of practice, in connexion with the function of nervous organs, we mean precisely that certain processes of excitation are facilitated: a result that can be produced most directly by an enhancement of excitability within the nerve-paths which the excitation travels. In saying this, we must, however, remember that the facts in question are facts of direct practice: that is, we must abstract from all the effects which practice can produce in other tissues, -- muscles, joints, tendons, bones, -- but which always make their appearance after a considerable interval' through these, in their gradual [p. 77] summation, constitute, of course, a very important part of the phenomena included in the usual definition of practice.
Suppose, however, that we allow the muscle to make the contractions which are the natural consequence of the stimuli applied to the nerve. We then invariably meet, after a certain lapse of time, with another phenomenon, which compensates the elementary phenomenon of practice described just now, and which presently reverses all the features of the picture. This is the phenomenon of fatigue. We can, therefore, observe both processes, practice and fatigue, in their simplest typical sequence, by making a muscle do work upon a weight of moderate size, which it has to lift, and by applying the stimuli at the appropriate intervals, with a brief interlude between twitch and twitch. Under these conditions, we have, first of all the effects of practice; the functional capacity of the nerve increases, quickly at the beginning, then more slowly. Then, from a certain point onwards, the height of lift remains the same, while the duration of the contraction is quite considerably increased. After a little while, however, the height of lift decreases, and the contraction is more and more prolonged. Finally, a single stimulus shock releases a weak but very slow contraction, -- similar to that provoked in the fresh muscle by the direct application of a constant galvanic current to the muscle-substance or, most markedly, by the passing of such a current through a muscle whose nervous excitability has been destroyed by curare poisoning.[11] The general character of these phenomena makes it probable that they have their principal seat, not in the nerve, but in the muscular substance itself. This hypothesis is, as a matter of fact, borne out by a number of different observations upon the phenomena of fatigue, which prove that they constitute, in this regard, a direct antithesis to the elementary phenomena of practice as described above. The latter can he obtained even when the muscle is entirely inactive; indeed, it is in such circumstances that they appear at their best. The fatigue phenomena, on the other hand, refuse to show themselves, so long as means are taken to prevent the contraction of the muscle during the application of stimuli to the nerve. With a sufficiently overloaded muscle, e.g., no amount of repeated stimulation win bring ot the signs of fatigue. If, in this case, a test-stimulus is applied to the nerve before and after the overloading of the muscle, the resulting contraction is just the same. Similar observations have been made upon animals temporarily deprived of the use of their muscles by poisons like curare or atropin, which paralyse the terminal apparatus of the motor nerves in muscle, but leave the nerve-trunks intact. If stimuli are applied to a nerve, during the action of the poison, there is no indication of nervous fatigue.[12][p. 78]
We must conclude from these results that the elementary phenomena of practice and fatigue are of radically different origin. The prime condition of the processes of practice is given in the nerve-substance, which is so constituted as to be very readily changed by stimulation: the change manifesting itself in a continuously increasing effectiveness of subsequent stimuli. All direct practice may be referred to this elementary phenomenon. Where it is shown by muscle, we may, in all probability, ascribe it to the nerves which the muscle contains, or to certain fundamental properties of contractile substance which nerve and muscle possess in common. With indirect practice, which appears as the result of actual, more especially of repeated exercise of function, the case is different. Here, we must admit, muscle plays a leading part: the increase of blood-supply, due to frequent repetition of contractions, means a more adequate nutrition, and consequently a higher functional capacity. These indirect effects of practice do not, however, differ in any essential respect from the changes produced in tendons, joints, bones, -- increased extensibility of tendons, smoothing of articular surfaces, etc., -- by frequent repetition of the same movement. They are secondary phenomenon, sharply marked off from the primary by the fact that they arise only by the mediation of changes in the blood-supply. On the other hand, the phenomena of fatigue resulting from the performance of mechanical work are as characteristic of muscle as the phenomena of practice are of nerve: they reside almost exclusively in the muscle-substance. And a like statement applies, by all analogy, to the other organs appended to the nervous system, the sense-organs and glands. Nervous substance itself seems to be, in large measure, exempt from fatigue. To explain this peculiarity, we must assume that it contains regulatory mechanisms, of a high degree of perfection, whose office is to prevent exhaustion. We have already spoken of the alternations of excitatory and inhibitory forces, evinced during the progress even of a simple process of excitation. It is these, then, we may suppose, that underlie the phenomenon of nervous inexhaustibility, guaranteeing a long period of functional activity, and protecting the nervous substance against injury of all kinds. At the same time, the inexhaustibility is, of course, a relative matter. There is good evidence that the effects of nervous exhaustion, when once it has set in, are all the more permanent, and that recuperation is all the more difficult. In view of this fact, the comparatively rapid fatigue of the peripheral organs appears in some sort as a measure of defence; it prevents any destructive consumption of nervous forces, by throwing the external instruments of nervous activity out of function before the nerves themselves are affected. [p. 79]
(d) -- Stimulation of Nerve by the Galvanic Current
We must now devote a special paragraph to the stimulation of nerve by the constant galvanic current. We are trying to gain an idea of the processes in nerve from a consideration of the course of stimulation-phenomena at large; and the phenomena which accompany this mode of stimulation will help us to fill in various details of the picture.
In general, the galvanic current affects the nerve by way of excitation, both at make and at break; but in both cases the processes of stimulation at anode and cathode are markedly different. With currents of not excessive intensity, the processes that follow directly upon the make of the current in the neighbourhood of the cathode are of the same character as those occurring after momentary stimulation throughout the whole length of the nerve; the only difference is that the excitatory and inhibitory effects persist, with diminished intensity, so long as the current is kept closed, while at the same time the excitatory processes remain constantly in the ascendant. In the neighbourhood of the anode, on the other hand, inhibitory forces of considerable intensity make their appearance. They increase, with increasing intensity of current, far more quickly than the excitatory effects; so that with fairly strong currents, when the anode lies nearer the muscle, the inhibition there set up prevents the propagation to the muscle of the excitation beginning at the cathode. The result is that, with increased intensity of the ascending current, the making contraction very soon decreases again, and presently disappears altogether. The anodal inhibition begins at the anode as soon as the current is made, and then diffuses slowly and with gradually diminishing intensity to a considerable distance. Its rate of travel, varying with the intensity of the current, is not more than 80 to 100 mm. in the 1 sec., -- very much slower than the rate of the excitatory process, which moves with a rapidity of 26 to 32 m. It should be noted, however, that this rate increases markedly with increase in the intensity of the current, so that the inhibition finally extends into the region of the cathode. If the current is now broken, the differences present during make disappear, more or less quickly, and at the same time inhibitory effects gain a temporary ascendency at the cathode; the break-stimulation thus consists in a process of compensation. It proceeds mainly from the region of the anode, where the inhibition maintained during make is transformed into excitation, -- the reversal occurring the more quickly, the stronger the current employed.
The peculiar features of the stimulation-processes released by the constant current may, then, be stated in summary as follows. The excitatory and inhibitory effects, which with other modes of stimulation are distributed uniformly throughout the nerve, here vary with the position of the electrodes: at make, the excitatory forces predominate in the neighbourhood [p. 80] of the cathode, the inhibitory in the neighbourhood of the anode; at break, a process of compensation sets in, which for a time exactly reverses the distribution of the two classes of effects.[13]
The phenomena of nervous stimulation are attended by other phenomena besides that of muscular work. Only the thermal and electrical changes, however, have so far been worked out in any detail. We may appeal to these to supplement, perhaps in some measure to check, the conclusions we have drawn from the phenomena of irritability. But we find, as was indicated above (p. 57) that the tale of results is exceedingly meagre. No one has as yet been able to demonstrate the occurrence of thermal changes in the nerve itself, in consequence of stimulation: but this simply means, of course, that the changes are too slight to be taken account of by our measuring instruments. On the other hand, heat is always set free when work is done by muscle, while at the same time the relation between development of heat and amount of mechanical work varies as the principle of the conservation of energy requires, -- increase of mechanical energy involving decrease of the relative quantity of heat developed. This fact comes out clearly, if we so arrange an experiment that the muscle shall make maximal contractions of equal height, while lifting weights of different sizes; the greater the weight to be raised, the smaller is the amount of heat generated.[14] In contradistinction to these differences in thermal phenomena, the electrical changes accompanying the process of stimulation in nerve and muscle have been shown to be alike. In both tissues, the point of excitation always becomes negatively electrical to any other unstimulated part. These changes cannot, however, be brought at present into any intimate relation with the processes of stimulation; our knowledge of the chemical conditions upon which they depend is glaringly defective. That a certain relation exists is shown, however, by their temporal course; the rate of propagation of the current of action in the nerve-fibre coincides with the rate of propagation of the stimulation-process itself. This coincidence extends, further, to the transmission of the inhibitory processes set up by the constant current, as described above; the changes occurring at the anode also travel much more slowly than those occurring at the cathode. The latter proceed with the same rapidity as the stimulus-wave at the rate of some 32 m. in the 1 sec.; the anodal wave of inhibition travels, as BERNSTEIN found, at the rate of only 8 to 9 m. in the 1 sec.[15]
§3. Theory of Nervous Excitation
The molecular state, which our general ideas of the mechanics of complex chemical processes lead us to predicate of nervous substance, was described above as a state in which there is constant performance at one and the same time of positive and negative molecular work. The positive molecular work, so soon as it gains the upper hand, will manifest itself either in disengagement of heat or in some form of external work, such as muscular con-[p. 81]traction. The negative molecular work will counteract these positive effects: heat will become latent; the progress of a muscular contraction will be inhibited. Equilibrium of the two opposed kinds of molecular work brings about the stationary condition of the nerve, during which there is no change of temperature and no accomplishment of external work. Hence, whenever we find that the action of an external stimulus releases a process which gives rise to a muscular contraction, or, for that matter, simply induces an increased irritability in presence of the test-stimulus, we may argue to an enhancement of the positive molecular work. Whenever, on the contrary, the progress of a muscular contraction is arrested, or the reaction to the test-stimulus reduced, we may be sure that the negative molecular work is in the ascendant. Whether, now, the one or the other of these effects is produced, whether, that is, the positive or the negative molecular work gains the upper hand, depends upon circumstances. We are thus led to the general conclusion that the stimulus shock increases both the positive and the negative molecular work of the nerve. This means, in terms of our preceding discussions, that the stimulus shock does two things: it assists the atoms of complex chemical molecules to unite in more stable connexions, while it also favours the disruption of these compounds and the return of the atoms to less stable and more complex relations. The restitution of the complex molecules corresponds to the recuperation of the nerve; the process of combustion, which ends in the formation of more stable and less readily decomposable compounds, is the source of the work which it performs, but is also the condition of its exhaustion. The only way in which the stimulus can bring about external work (muscular contraction the excitation of nerve-cells) is by furthering the positive molecular work more effectively than the negative. The positive molecular work then becomes the source of the external work of excitation, which may he transmitted to particular organs, and so still farther transformed into other modes of work. At the same time, the positive and negative molecular work must be distributed over the course of stimulation in the sequence determined by the relation of the excitatory to the inhibitory effects. First of all, that is, there must be a storing-up of potential work, corresponding to the stage of inexcitability; the stimulus shock releases a number of molecules from their existing connexion. Thereupon begins a process of combustion, starting with the freed particles, and extending from them to the readily combustible constituents of the nerve-mass at large; during this stage a large quantity of potential is transformed into actual work. If the combustion proceeds with great rapidity, it is followed for a short time by a restitution of the complex molecules (transitory inhibitions; preponderance of negative molecular work). As a rule, however, there remains at the conclusion of the contraction a surplus of positive molecular work, which disappears only gradually; we trace it [p. 82] in the enhanced effect of a second stimulus supervening upon the first. It follows, then, that the same curves which we employed to illustrate the relations of excitation and inhibition (Fig. 30, p. 75) will serve here to show the relation of positive to negative molecular work. The equilibrium of the two, during the state of rest, is indicated by the equality of the initial and terminal ordinates, x a, x c and x b, x d. We must, however, suppose that the internal condition of the nerve, after the process of stimulation has run its course, is not in general precisely the same as before: there will, on the whole, have been more given out in positive work than has been acquired in negative in potential work. Nevertheless, we must also infer, from the fact of the relative inexhaustibility of nerve, that this difference is extremely small; so that the equilibrium of forces is re-established in large measure: and in nerves of high functional capacity probably in full measure, during the actual progress of the contraction. This tendency to the maintenance of equilibrium between positive and negative molecular work, between loss of work-equivalents and gain of potential work, appears to be a specific property of nervous substance, founded in its chemical constitution and distinguishing it from all other tissues. It is expressed symbolically in Fig. 30, which shows the molecular processes of stimulation; the upper and lower curves each include an approximately equal area. This implies that the process of stimulation consists essentially not in a permanent disturbance of the equilibrium between positive and negative molecular work, but simply in their different distribution in time during the progress of the stimulation. The nature of this difference is given at once with the changes of irritability that can be traced from moment to moment in the figure.
We must, now, not lose sight of the fact that it is never more than a certain position of the total sum of positive molecular work, set free in the nerve by stimulation, that is transformed into excitatory effects, or, as we may phrase it, into work of excitation: another portion may become heat, a third be changed back again into potential (negative) work. Similarly, it is only a portion of the work of excitation that is employed in the production of external stimulus effects (muscular contraction or stimulation of nerve-cells); we have seen that there is always an enhancement of irritability, both during and after the contraction. Hence a supervening stimulus will invariably and the nerve possessed of a surplus of work of excitation. If no new stimulus shock supervenes, this surplus in all probability passes over into heat. After the work of excitation has once been set up, at the point of stimulation, it exerts an influence upon neighbouring parts, where the store of molecular work is in its turn transformed in part into work of excitation, and so on. But, as we know, the process released by the momentary stimulus persists for a considerable time. Hence, while work [p. 83] of excitation is released, new stimulus-impulses are conveyed to the part affected from the neighbouring parts. We are in this way able to explain the heightening of excitation observable when different points of the nerve are subjected to stimulation (p. 68).
The main difference between these general stimulation-processes and stimulation by the constant current is, obviously, to be found in the uneven distribution of the sums of positive and negative molecular work which obtains in the latter case. While the current is closed, there is preponderance of negative molecular work in the neighbourhood of the anode, of positive in the region of the cathode. This difference becomes intelligible, when we remember that the resulting electrolysis must produce internal changes in the nerve-substance. At the positive electrode electronegative, at the negative electrode electropositive constituents are given off. At both places, that is, the word of the electric current produces dissociation. The immediate consequence is, that work must disappear; but as soon as the wandering partial molecules tend to enter into more stable compounds than those from which they have been separated, the positive molecular work may begin to increase, that is, a part of the work which has disappeared may be set free again. The phenomena of stimulation lead us to infer that the first of these processes takes place regularly in the neighbourhood of the cathode, the second in the region of the anode. The precise chemical changes involved are as yet unknown to us; but the phenomena of electrolysis supply an abundance of analogous instances of the interchange of forces. Thus, in the electrolysis of stannous chloride, we have at the cathode a deposition of tin, in which the work employed for its separation remains stored as potential work, while at the anode we obtain chlorine, which at once unites with the stannous chloride to form stannic chloride, liberating heat in the process. Similar results may appear in all cases where the products of electrolysis are liable to chemical interaction. At break of a current passing through a length of nerve, on the other hand, a less well-marked process of electrolytic decomposition sets in, as a consequence of its polarisation, in a direction opposed to that of the original current. This, together with the gradual compensation of the chemical differences, occasions the phenomena of the break-stimulation.
We may say a word, in conclusion, of the relation of the processes whose general mechanism we have here described to the electrical changes in the stimulated nerve. It is a noteworthy fact that the current of action which follows upon a momentary stimulation of the nerve reaches its conclusion, on the average, as early as 0.0006 to 0.0007 sec. after the application of the stimulus,[16] and therefore falls completely within the period of nervous in-[p. 84]excitability.[17] It would appear, then, that the variation is connected with the inhibitory forces, or with the passage of positive into negative molecular work. We must, however, have further information, as regards the character of this connexion, before we can think of turning the electrical processes to theoretical account.
The phrases 'positive' and 'negative molecular work' are meant to suggest the general line of thought followed by the science of mechanical energy or, as we may put it more briefly, by mechanical energetics. Modern physiologists not infrequently substitute for them the words 'assimilation' and 'dissimilation,' antithetical terms borrowed from the vocabulary of metabolic phenomena, and thence transferred to that of the general mechanics of the nervous system. It is, the author hopes, hardly necessary to point out in this place that the words and phrases employed in the text are not translations back again from the language of physiological chemistry into that of mechanics; though in such an idea, might possibly arise, in view of the popularity of the antithesis 'assimilation-dissimilation' at the present time and the variety of contexts in which it appears. The contrary is true: the phraseology adopted in the text, and modelled upon that of a general mechanical energetics, was followed in the first edition of this work (1874) and in the still earlier "Untersuchungen zur Mechanik der Nerven" (i., 1871), before the terms 'assimilation' and 'dissimilation' had begun to play their part in physiology. Its retention in the present edition is not due to any prejudice on the author's part in favour of the original form in which his thought was cast, but rather to the objections which he feels may be urged against the alternative wording. The processes of metabolism which, for want of more precise terms, we name tentatively 'assimilation' and 'dissimilation' are, apart from the effects from which the names are derived, altogether unknown to us; and the effects themselves are simply that, in the one case, an existing tissue is reinforced by complex tissue-materials of the same order, while, in the other, existing tissue-materials are brought to disappearance. We have good ground for the assumption that, in dissimilation, the decomposition of the complex molecules, and the combustion-processes resulting from it, have the upper hand; and we may suppose that in assimilation, conversely, the chemical processes introduced are predominantly synthetic in character. But when we ask how in detail the interchange of nerve is effected in the two cases, the reply is that as regards dissimilation very little, and as regards assimilation practically nothing is known. The various processes involved certainly do not conform to any simple pattern, but depend upon a series of chemical interactions so complicated that at present we have no means of tracing them. This complication is vouched for by the fact, now fairly well established for all such processes, that in dissimilation there is a constant interplay of decomposition and recomposition; existing chemical connexions are dissolved, and new compounds formed, in continual interdependence. As a rule, energy is liberated during these dissimilations, in the form of heat or of mechanical work. But again, we cannot say which phase of the dissimulation process is responsible for this result; nor do we know if a similar interchange [p. 85] of energy is necessarily involved in every instance of dissimilation, or if there may not be processes, whose chemical effects would lead us to class them with the others as dissimilations, bat whose general and final effect is accompanied by a transformation of energy in the opposite sense. Our knowledge of the chemism of the metabolic processes is, in the author's opinion, far too defective to permit of our answering these questions. The theory of assimilation and dissimilation thus attempts to illuminate the processes in the nervous system by analogies that are more obscure than the processes themselves; and such a proceeding can hardly inspire confidence. It is sometimes said that dissimilation is the correlate of fatigue, and assimilation of recuperation. But to say this is, after all, merely to set in place of certain complex symptoms conditions that are at least as complex, and far less open to demonstration.[18] Fatigue and recuperation are symptomatic terms whose meaning is, roughly at any rate, clear to everyone. It is, as we have seen, highly probable that the metabolic processes which give rise to both groups of symptoms are extremely complicated. When we consider, therefore, that the words 'assimilation' and 'dissimilation' used to denote these processes are words that have no chemical significance, but are in the last resort purely teleological concepts, we cannot but suspect that the symptomatic terms 'fatigue' and 'recuperation' are simply coming back to us in changed form. If we wish to analyse these latter in detail, there are, as it seems to the author, but two ways open, in the present state of our knowledge. We may, on the one hand, limit ourselves to the symptoms, but, while we do this, attempt so far as may be to reduce the phenomena, given in an extremely complicated syndromes, to their simplest components. We thus discover that every process of fatigue and recuperation contains two elementary terms, variously interrelate in the particular case, -- excitation and inhibition. Or, on the other hand, we may attempt to refer the effects in question to the more general concepts furnished by mechanical energetics. We then arrive at the notion of positive and negative molecular work, in the sense in which these phrases have been employed in the text. This restriction of the hypothetical foundations of nerve physiology to straight forward physical analogies appears to the author to be especially desirable, in view of the great extension of the rival terms. We find the antithesis of 'assimilation' and 'dissimilation' in the most diverse contexts, -- in the theory of visual sensations, in the theory of auditory sensations, applied to all other conceivable physiological and psychophysical phenomena. It almost seems, indeed, as if this pairs of terms is gradually coming to play the part in modern physiology that was played in SCHELLING'S nature-philosophy, at the beginning of the nineteenth century, by the phrase "polar opposites," which found application not only to electricity, magnetism and chemical process, but also to sensibility and irritability, light and darkness, and many other things besides.
§4. Influence of the Central Parts upon the Processes of Excitation
(a) -- Course of the Reflex Excitation
We begin our investigation of the processes in the central nervous substance by stimulating a peripheral nerve, and endeavouring to find out how the course of stimulation is altered, if it is compelled to pass through central [p. 86] elements. The easiest way to perform this experiment is to avail ourselves of the phenomenon of reflex excitation. We first of all apply an electric shock of the proper intensity to a motor nerve root, whose connexion with the myel on the one hand and with its dependent group of muscles on the other is kept intact; and we then stimulate, in the same way, the central end of some sensory root. The two twitches are recorded by the muscle, and the experiment is so arranged that the times of stimulation correspond to the same point upon the axis of abscissas of the two muscle-curves. The differences in appearance and progress of the two contractions then give us a measure of the influence exerted by the intercalated central substance.
The first thing that we observe, under these circumstances, is that much stronger stimuli are required to produce contraction by way of the sensory root. If we make our shocks as nearly instantaneous as may be, e.g. by using induction-shocks, we shall frequently find it impossible to release any reflex contraction whatsoever; the currents required are of such intensity that their employment would bring with it the danger of leakage to the myel.[19] Provided, however, that the reflex irritability is high enough to permit of our making the experiment, we obtain two curves which repeat, on a greatly enlarged scale, the same characteristics that distinguish the curves obtained in a previous experiment by stimulation of a motor nerve at two points unequally distant from the muscle (cf. Fig. 24, p. 67) The reflex twitch is extraordinarily late in appearing, and is of much longer duration. Suppose, e.g., that we stimulate a motor and a sensory root which enter the myel at the same height and on the same side, and that we so regulate the stimuli as to equalise the heights of the muscle curves; we get the result shown in Fig. 31. The only marked difference between these contractions and those released from different points upon the motor nerve is that a  stronger and not a weaker stimulus must be employed, to bring the reflex contraction to the same height as the other. The differences in the course of the excitation are, however, in this instance so considerable, that no increase in the intensity of stimulus is able to change their character. It is true that intensification of stimulus increases not only the height but also the duration of the contractions, while decreasing the latent period. But [p. 87] the weakest reflex contractions are always noticeable by their long duration, and the strongest by the length of their latent period, even when we compare the former with the strongest and the latter with the weakest direct contractions.[20] It is clear, now, that the time which the stimulation requires to pass from a sensory into a motor root is given by the interval separating the beginnings of the two contractions, the direct and the reflex. The nerve-roots are so short, that the portion of this interval taken up by the peripheral conduction may be considered negligible; and we may accordingly designate the interval as a whole, the reflex time. To determine it, we must -- since the latent period is dependent upon the intensity of the stimuli -- have recourse once more, as we did in our measurement of the rate of propagation in the nerves, to experiments in which the muscle-nerves are of the same height.
stronger and not a weaker stimulus must be employed, to bring the reflex contraction to the same height as the other. The differences in the course of the excitation are, however, in this instance so considerable, that no increase in the intensity of stimulus is able to change their character. It is true that intensification of stimulus increases not only the height but also the duration of the contractions, while decreasing the latent period. But [p. 87] the weakest reflex contractions are always noticeable by their long duration, and the strongest by the length of their latent period, even when we compare the former with the strongest and the latter with the weakest direct contractions.[20] It is clear, now, that the time which the stimulation requires to pass from a sensory into a motor root is given by the interval separating the beginnings of the two contractions, the direct and the reflex. The nerve-roots are so short, that the portion of this interval taken up by the peripheral conduction may be considered negligible; and we may accordingly designate the interval as a whole, the reflex time. To determine it, we must -- since the latent period is dependent upon the intensity of the stimuli -- have recourse once more, as we did in our measurement of the rate of propagation in the nerves, to experiments in which the muscle-nerves are of the same height.
This presupposed, we may proceed to investigate the reflex time under various conditions. The simplest case is shown in Fig. 31, where the stimulation is transferred from a sensory root to a motor root belonging to the same nerve-trunk: we may term this the case of same-sided reflex excitation. Next in order comes the propagation of stimulus from a sensory root to a motor root which leaves the myel at the same height but upon the opposite side: we term this the case of crossed reflex excitation. In the third place, we may have propagation along the length of the myel, which we may call the longitudinal conduction of reflexes; as, e.g., in the transference from the sensory root of a nerve of the arm to the motor root of a nerve of the lower extremities. In no one of these three cases is the reflex time sensibly dependent upon the intensity of the excitations. It is, as might have been predicted, relatively shortest for same-sided reflex excitation, where under normal circumstances it amounts to 0.008 to 0.015 sec. It is, however, as one would be less likely to expect, relatively greater with crossed than with longitudinal conduction. Thus, if we compare the crossed with the same-sided reflex, there is an average difference to the disadvantage of the former of some 0.004 sec. If we then compare the reflex released in the thigh by stimulation of the root of a sensory nerve of the arm with the same-sided reflex, the difference between the two times is as a rule somewhat smaller.[21] Since the path travelled by the stimulation in the latter case is at least six to eight times as long as that traversed in the former, it is evident that the retardation in crossed conduction is much more serious than it is in longitudinal conduction. An explanation is, without any doubt, to be found in the fact that longitudinal conduction (as we shall see presently, in Chapter V., when we come to discuss the morphology of the myel) is sub-[p. 88]served for the most part by the fibres of the white substance, while crossed conduction must be mediated almost exclusively by the cell-reticulum of the grey substance. We have, then, in the results of this set of experiments, a confirmation of the inference, already suggested as probable by the long duration of the reflex time, that the central elements offer incomparably more resistance than the nerve-fibres to the progress of an excitation. The same conclusion may be drawn from the further fact that a retardation of conduction, amounting on the average to 0.003 sec., occurs in the spinal ganglia of the frog, and again from the related observation that the sensory nerve roots are more irritable than the nerve-fibres below the spinal ganglia. It is noteworthy, in connexion with this latter result, that the ramifications of the sensory nerves in the skin are, in their turn, more easily excitable than the nerve-branches that run to the skin. Just, then, as there are mechanisms in the spinal ganglia which diminish the irritability of the incoming nerves, so must there he mechanisms in the skin which discharge a precisely opposite function. It follows from all this that the irritability of the nerve-trunks and their branches is reduced to a minimum: a characteristic, we need hardly say, that is eminently fitted to protect the central organs from the advent of useless sensory excitations.[22]
(b) -- Enhancement of Reflex Excitability
The temporal relations of reflex conduction have made it appear probable that the central elements, while on the one hand they offer greater resistance to incoming excitations, are able, on the other, to develop a greater amount of stored energy. This hypothesis is confirmed by many other facts. We notice, first of all that in almost every case, where the excitability of the myel is not enhanced by artificial means,[23] a single, momentary stimulus-shock is unable to release a reflex contraction. To obtain a response, we must repeat the stimulation; and the contraction thus set up usually takes on a tetanic character.[24] Within certain limits, the reflex makes its appearance after the same number of single stimulations, whether these be given in quick or slow succession. On the other hand, the duration of a [p. 89] reflex tetanus is not directly dependent upon the duration of the stimulation, as is that of the contraction aroused by tetanic excitation of the motor-nerve. If the stimulation be of short duration, the tetanus outlasts it; if it be of longer duration, the tetanus disappears earlier than the stimulation itself.[25] Another phenomenon, that shows very clearly the differences in excitability between the peripheral and the central nervous substance, is the following. If we stimulate the motor nerve by induction-shocks, repeated with not too great rapidity, the corresponding muscle falls, as was first pointed out by HELMHOLTZ [26] into vibrations of the same frequency. These may be perceived as a tone, or may be recorded, by means of a fitting instrument, upon a cylinder which rotates with uniform speed. If, now, we  stimulate the myel in the same way, the muscle again falls into vibrations, But the frequency of vibration is considerably diminished. Fig. 32 shows two curves of vibration obtained from the muscle of a rabbit by KRONECKER and HALL. With 42 stimuli in the 1 sec., the muscle traced the upper curve when the motor nerve was stimulated, the lower, when the stimulation was applied to the myel, which had been severed below the oblongata. [27] Closely connected with this is BAXT'S observation that voluntary movements, however simple they may be made, always last a considerably longer time than simple contractions, released by the stimulation of a motor nerve. BAXT found, e.g., in experiments upon himself, that the index finger of the right hand, moving in response to stimulation by the induction current, required on the average 0.166 sec., while movement initiated by voluntary innervation required 0.296 sec.[28]
stimulate the myel in the same way, the muscle again falls into vibrations, But the frequency of vibration is considerably diminished. Fig. 32 shows two curves of vibration obtained from the muscle of a rabbit by KRONECKER and HALL. With 42 stimuli in the 1 sec., the muscle traced the upper curve when the motor nerve was stimulated, the lower, when the stimulation was applied to the myel, which had been severed below the oblongata. [27] Closely connected with this is BAXT'S observation that voluntary movements, however simple they may be made, always last a considerably longer time than simple contractions, released by the stimulation of a motor nerve. BAXT found, e.g., in experiments upon himself, that the index finger of the right hand, moving in response to stimulation by the induction current, required on the average 0.166 sec., while movement initiated by voluntary innervation required 0.296 sec.[28]
We can easily see the reason for the greater effect produced upon the myel by frequent repetition of the stimulus. Every stimulation leaves behind it an enhanced reflex excitability. Here, again, however, the central substance merely exhibits, on a larger scale, phenomena with which we are already familiar in the case of the peripheral nerve. On the other hand, there are certain chemical effects, that are able in some unknown way to produce a similar change of irritability, which appear to be peculiar to the central nerve-substance. The agents in these effects are termed 'reflex poisons'; the chief place among them is taken by strychnine, which brings abort the changes in question with unfailing certainty. Strychnine prob-[p. 90]ably owes its power in this regard to the circumstance that its effect is limited almost exclusively to the central substance of the myel; whereas other nerve-poisons set up changes in the peripheral nerves, or in higher nerve-centres, which may serve, in greater or less degree, to counteract the effect under discussion.[29]
The action of these poisons is, in general, as follows. (1) Much weaker stimuli are sufficient to release a reflex contraction; indeed, a point is very soon reached, at which the reflex irritability becomes greater than the irritability of the motor nerve. (2) Even when the stimuli are reduced to the lowest limit of effectiveness, the contraction is higher and, in particular, of longer duration than under normal conditions; if time effect of the poison be increased, it passes over into a tetanic contraction. (3) The beginning of the contraction is more and more delayed; so that the latent period may have more than twice its ordinary duration. At the same time, the difference in the length of the latent period with strong and weak stimuli is enormously increased: when the 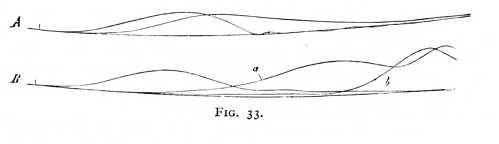 action of the poison is at its height, the reflex tetanus hardly shows any difference of degree, whether the weakest or the strongest stimuli be applied, but in the former case sets in with extraordinary slowness. An illustration of these changes is given in Fig. 33. The curve A was taken as the action of the poison was beginning; the curves B, when it was at its full height: a was released by a strong, b, by a weak momentary stimulus: in both cases a direct contraction has been recorded, for purposes of comparison. There can be no doubt that this lengthening of the latent period is directly connected with the enhancement of irritability. When the central substance is modified by the poison, the after-effect of the stimulus is prolonged, so that the excitation call be released after the initial inhibition has been overcome. The phenomenon is somewhat like that of the summation of stimulations, only that here the external stimulus is not repeated. We must, accordingly, suppose that the stimulus brings about a number of successive stimulations, whose summation presently leads to excitation. This suggests the idea that the processes of molecular inhibition are not sensibly changed by the alteration of the [p. 91] nervous substance, but that the positive molecular work is seriously affected. In the normal state, it returns to the potential form, in whole or in great part, immediately after its liberation; in the present instance, it appears to be recovered but gradually. We may note that similar, though weaker, effects are produced on the myel by the action of cold.[30]
action of the poison is at its height, the reflex tetanus hardly shows any difference of degree, whether the weakest or the strongest stimuli be applied, but in the former case sets in with extraordinary slowness. An illustration of these changes is given in Fig. 33. The curve A was taken as the action of the poison was beginning; the curves B, when it was at its full height: a was released by a strong, b, by a weak momentary stimulus: in both cases a direct contraction has been recorded, for purposes of comparison. There can be no doubt that this lengthening of the latent period is directly connected with the enhancement of irritability. When the central substance is modified by the poison, the after-effect of the stimulus is prolonged, so that the excitation call be released after the initial inhibition has been overcome. The phenomenon is somewhat like that of the summation of stimulations, only that here the external stimulus is not repeated. We must, accordingly, suppose that the stimulus brings about a number of successive stimulations, whose summation presently leads to excitation. This suggests the idea that the processes of molecular inhibition are not sensibly changed by the alteration of the [p. 91] nervous substance, but that the positive molecular work is seriously affected. In the normal state, it returns to the potential form, in whole or in great part, immediately after its liberation; in the present instance, it appears to be recovered but gradually. We may note that similar, though weaker, effects are produced on the myel by the action of cold.[30]
(c) -- Inhibitions of Reflexes by Interference of Stimuli
We have spoken so far only of the influences which enhance the excitability of the central elements. Here, however, just as in the case of the nerve-fibre, the excitatory are paralleled by inhibitory effects. The fact that first drew attention to these inhibitions is a discovery of old standing in physiology: the fact that the reflex excitability of the myel is increased after removal of the brain. SETSCHENOW, starting out from this fact, found that the stimulation of certain parts of the brain in the frog, -- thalamus, bigemina, medulla, --prevents or delays the appearance of the reflexes.[31] He was therefore inclined to think that the inhibitory function is confined to certain definite central parts. Further experiments showed, however, that the same effect is produced by the stimulation of other sensory nerves, or of the sensory columns of the myel; [32] so that it became necessary, in terms of SETSCHENOW'S hypothesis, to suppose that these specific inhibitory centres are distributed over almost the entire cerebrospinal organ. But if any given sensory excitation may be inhibited by the stimulation of any other sensory element, the sphere of inhibition -- as GOLTZ justly observed [33] becomes coextensive with the sphere of sensory excitation; and the assumption of specific inhibitory centres falls to the ground. At the same time, while any possible sort of sensory stimulation, whether it affect other sensory nerves or sensory central parts, may arrest the progress of a reflex [p. 92] excitation, the inhibitory effect is not by any means invariably produced; the supervening stimulation may, on the contrary, enhance the reflex, -- as always happens, of course, when two excitations meet in some motor fibre, or in a motor central area. Let us term the meeting of two excitations in the same central territory, quite generally, an interference of stimulations. Then the result of such interference is dependent upon four things. It depends (1) upon the phase to which the one excitation has attained when the other begins. If the muscle contraction released by the first stimulation is still in course, or only just over, when the second arrives, we have as a rule an enhancement of the stimulus-effect. If, on the other hand, the original stimulation occurred some time before the application of the second, this latter is more easily inhibited. It depends (2) upon the intensity of the stimuli. Strong interference-stimuli inhibit a given reflex excitation more easily than weak; sometimes, indeed, strong stimuli will inhibit the same excitation that weak stimuli enhance. It depends (3) upon the spatial relation of the nerve-fibres stimulated. Sensory fibres that enter the myel at the same height and upon the same side, i.e. that belong originally to one and the same nerve-trunk, effect a much weaker inhibition (or, in other terms, are much more ready to enhance the excitation) than fibres that come in from different sides or at different levels. Lastly, it depends (4) upon the state of the central organ. The more completely the normal functional capacity is preserved, the more certainly, other conditions being favourable, may one look for inhibition of the reflexes ; the more the functions of the organ have been impaired by cold, by strychnine or other reflex poison, by loss of nervous force due to fatigue, malnutrition, etc., the more likely is it that an enhancement of stimulation will take the place of inhibition. This decrease of inhibition is evidenced, first of all by the fact that stimuli of longer duration and greater intensity are required to evoke it. It always disappears first with stimulation of the nerve-fibres belonging to the same root; but in a state of extreme functional incapacity, or of serious derangement by cold or strychnine, it disappears altogether, so that no inhibitory symptoms can he observed at all.[34]
It is, perhaps, tempting to think of these inhibitory effects due to an interference of oscillatory stimulus-motions, analogous to the interference of light and sound vibrations; to conceive, i.e., that the stimulus-waves meet together and, in whole or part, cancel one another.[35] Such an hypothe-[p. 93]sis is, however, wholly unable to explain the simple effacement of excitation that occurs, e. g., in the ventral nerve-cells of the myel, when the motor fibres issuing from them are stimulated. Moreover, it gains no support from the known facts of the course of excitation. On the contrary, the varying results of stimulus-interference indicate, quite clearly, that in the stimulation of the central elements, as in that of the nerve-fibre, excitatory and inhibitory effects are released at one and the same time. It is also clear, however, that the phenomena of inhibition are in this case much more pronounced than they are in the peripheral nerve-fibre. The special conditions under which the two opposed results of central stimulation are obtained make it probable, further, that the external effect of inhibition is produced more particularly when the stimuli are so conducted as to interfere in the same sensory central area; whereas summation of stimuli seems to occur whenever the excitation travels from different sensory central areas, simultaneously stimulated, to the same motor elements. In general both of these effects may he produced, side by side, by the simultaneous stimulation of any different sensory elements, and it will depend upon the special circumstances whether the one or the other of them gains the upper hand.
(d) -- Chronic Effects of Excitation and Inhibition: Positive and Negative Tonus
If we inquire into the nature of these specific circumstances, we find as the most important, the connexions in which the various nervous elements stand with one another and with their appended organs. This conclusion is suggested at once by certain phenomena observed in nerves and muscles whose functional connexion with their central points of origin has remained intact. Thus, in the first place, a muscle which is united by its nerve to the central organ is kept permanently in a certain tension, which ceases at once, as may be observed in the slight lengthening of the loaded muscle, when the nerve is cut through.[36] This permanent tension in the state of rest is known as the tonus of a muscle. Its disappearance when the nerve is cut indicates that it has its ground in a chronic excitation of the nerve, transmitted to the fibre from the central elements. Its maintenance seems to depend, further, upon the connexions in which the central elements stand with one another. For the tonic excitation which travels to the skeletal muscles along the motor nerves of the myel may be abrogated, not only by section of the motor nerves themselves, but also by section of the sensory roots of the spinal nerves.[37] We must therefore [p. 94] suppose that a portion of the forces which release the excitation reach the motor nerve-cells only by way of their connexions with sensory elements; while the related observation, often made and confirmed, that tonus persists after the severance of such sensory connexions, points us to the cells of origin of the motor nerve-fibres as a co-ordinate and independent source of excitatory forces. On the other hand, however, the central elements appear also under certain conditions, according to the circumstances in which they are placed by their nearer or more remote connexion with other like elements, to generate, and to transmit to their peripheral continuations, a surplus of inhibitory forces. Here, e.g., belongs the observation that increase of tonus in a determinate muscle-group is regularly followed by decrease of tension in the antagonists: so that increased excitation of the flexor muscles of a limb brings with it a decrease of excitation in the extensors, and conversely. [38] We may term this phenomenon that of negative tonicity. It then becomes evident that the two opposing forms of tonus may be brought into relation with the fundamental phenomena of excitation and inhibition, which we have seen to be observable, first in the peripheral nerve, and then, on an enlarged scale, throughout the central organs of the nervous system. We have only to add, what is shown by all these observations, that the continual shift of government from excitation to inhibition, and back again, is very largely dependent upon the influences to which the central elements are subjected, in virtue of their connexion with other like elements and with the stimulation processes which these latter convey to them.
§5. Theory of Central Innervation
(a) -- General Theory of the Molecular Processes in the Nerve-Cell
The phenomena of central innervation have referred us to the same two classes of opposed molecular effects that we traced in the process of excitation in the nerve-fibre, The general view that we were led to take of this latter will, therefore, serve as our point of departure in the present instance. We begin, accordingly, by postulating for the central substance a stationary condition, similar to that which we assumed in the nerve-fibre; a condition, i.e., in which there is an equilibrium of positive and negative molecular work. The application of stimulus, here as before, means an increase in the amount of both forms of work. But everything points to the conclusion that, in the nerve-cell, there is at first a marked prepon-[p. 95]derance in the increase of the negative molecular work; so that a momentary stimulus-shock is, as a rule, unable to release any excitation whatever. If, however, the stimuli are repeated, then, as one follows another, the amount of negative molecular work is gradually diminished, in proportion to the positive, until at last this latter attains such dimensions that excitation arises. We may therefore suppose that the typical process of stimulation in a nerve-cell is analogous to that set up at the anode in a nerve-fibre by the making of a constant current. Under the action of the stimulus, the processes which transform more stable into less stable compounds, i.e. which subserve the storage of potential work, are thrown into increased activity. There is, however, a difference. When the current is applied to the nerve, its electrolytic action introduces decomposition-processes which do not normally take place in the nerve-fibre. When, on the other hand, the nerve-cell is stimulated, we have no right to assume anything more than an enhancement of activity which, under ordinary circumstances, is directed mainly upon the formation of complex chemical molecules, i.e. upon the accumulation of potential work. This difference between nerve-fibre and central substance, whose importance is sufficiently evident, is attested by other physiological considerations. The nerve-cells are really the laboratories, in which the materials that compose the nerve-mass are prepared. In the nerve-fibres, these materials are very largely consumed, in consequence of physiological function, but -- if we abstract from the inadequate and partial restitution which accompanies decomposition in every case of stimulation -- cannot, obviously, be reformed. For the fibres, if we separate them from their cells of origin, lose their nervous constituents, and the renewal of these proceeds always from the central points.[39] Even in the state of functional inactivity, therefore, the interchange of materials and forces within the nerve-cell is not in perfect equilibrium. But the balance dips on the one side in the cell, on the other in the fibre. Characteristic of the nerve is the formation of definite products of combustion, with the performance of positive work; characteristic of the cell is the production of compounds of high complexity, in which potential work is stored. It is true that the work done in the animal body, as a whole, is pre-eminently positive work, the combustion of the complex organic compounds; but it is altogether wrong to look upon this as the only means for the interchange of forces and materials within the organism. There are always going on, alongside of the positive work, reductions, dissolutions of more stable into less stable compounds, with the resulting accumulation of potential work. The nervous system, in particular, is the scene of great activity in this regard. The compounds which enter into the formation of nervous substance are, in some cases; more complex, [p. 96] and possessed of a higher combustion-value, than the nutritive materials from which they are derived; that is, are compounds in which a large amount of potential work is stored up.[40] The nerve-cells, the architects of these compounds, are in a certain sense akin to the plant-cells. These, too, accumulate potential work, which may remain latent until need arises, and then be transformed back again into actual work. The nerve-cells, in the same way, are the storehouses in which materials are laid up for future use. And the chief consumers of these stores are the peripheral nerves and their terminal organs.
Putting all this together, we may gain some idea of the relation obtaining between the central substance and the nerve-fibres that issue from it. We have, first of all the transmission from cell to fibre of those molecular motions that we term processes of excitation. But this is by no means all. There is, further, a constant movement of material, in the direction from centre to periphery; so that the fibre is in continual receipt of substances in which potential work is stored up. Here, it is plain, we have the explanation of the nutritive influence which the central substance everywhere exerts upon the nerve-fibres connected with it, and, through their mediation, upon the organs which they supply. This nutritive function belongs to all nerve-centres and nerve-fibres, and is intimately connected with the general mechanics of central innervation. The hypothesis that there is a special class of nerves, specially devoted to trophic functions, seems therefore to have nothing in its favour. The conditions under which this movement of material takes place must, however, necessarily react upon the phenomena of irritability and the course of excitation. Suppose, e.g., that a certain central area has enjoyed a long period of rest, and has consequently accumulated a large store of potential work. The actual work, sensory or motor, done in this area itself and in the nerve-fibres connected with it will, in general, be more intensive and of longer duration than would have been the case under different conditions. It is, also, not improbable that the movement of material may serve to develope neurodynamic interaction between adjoining central parts, as a result of which the actual work done at any given point may be increased by the conveyance of potential work from neighbouring points.[41] The differences in the response of the nerve-cells to the stimuli conducted to them proves, further, that every cell is divided into two distinct regions, the one of which resembles in excitability the peripheral nerve- substance, while the other shows a marked degree of divergence. We will term the former the peripheral, the latter the central region of the nerve-cell.[p. 97] The central region, we may suppose, is devoted pre-eminently to the formation of the complex compounds of which the nervous substance is composed; it is, therefore, the place of storage of potential work. A stimulus-movement conducted to it simply accelerates the molecular processes in the direction in which they are already moving, and accordingly disappears without external effect. It is different with the peripheral region. Here, too, something is done towards the transformation of actual into potential work. But, besides this, there is already a fairly rapid consumption of materials, derived in part from the central region, and a consequent production of work. If a stimulus strikes this peripheral region, its first result is, again, a relatively greater increase of the negative than of the positive molecular work. But the negative soon sinks back to its ordinary level, while the positive persists for a considerable time; so that, perhaps after an unusually long latent period, certainly if the original stimulus is reinforced by new stimulus-impulses, it is able to produce an excitation. For the rest, here as in the nerve, it is only a portion of the positive molecular work that passes over into work of excitation, and again only a portion of this that shows itself in external excitatory effects; another portion may be transformed back again into negative molecular work, and the work of excitation may be changed, in whole or part, into other forms of molecular motion. Further, when once excitation has arisen, the accumulated work of excitation is consumed very rapidly; the process suggests that of explosive decomposition. At the same time, the greater strength of the inhibition has meant the storage of a correspondingly greater amount of work of excitation; so that the stimulus-effect, when it appears, is greater than in the case of nerve-stimulation. In this respect, the irritable region of the nerve-cell stands to the peripheral nervous system in somewhat the same relation that a steam-boiler with stiffly working valve bears to a similar boiler whose valve moves easily. The expansive force of the steam must be made more considerable, in the former case, if the valve is to be opened; but, when this is done, the steam rushes out with a correspondingly greater force. It should be added that the peripheral region of the nerve-cell probably evinces a different conduct in different cases, approaching sometimes more, sometimes less nearly to the character of the peripheral nerve-substance. Thus, the sensory excitations conducted upward by the cells of the dorsal cornua of the myel are certainly less changed than the reflex excitations which are mediated as well by the cells of the ventral cornua. These differences may be conditioned upon the number of central cells which the stimulation has to traverse. But it is also conceivable that there is a continuous transition from the one to the other of the two cell-regions which we have named the central and the peripheral and that certain fibrils terminate in middle regions, in which inhibition is not yet [p. 98] complete, while at the same time difficulties are placed in the path of the stimulation.
We are now in a position to interpret the peculiar enhancement of reflex excitability produced by the repetition of stimulus or by the action of poisons. Under these conditions, the positive molecular work, once liberated, can be retransformed into negative work either not at all or, at least, less completely than usual. It therefore accumulates, until excitation arises. The effect of these two modes of interference is, therefore, to prevent the restitution of the nerve-substance, and so to make it possible for comparatively weak external impulses to set up a rapidly extending decomposition, in consequence of which the stored forces are soon exhausted.
There are still two things to be explained: the phenomenon of the mutual inhibition of excitations conveyed to the same nerve-cells from different quarters, and the fact that the stimulation can traverse certain cells only in one direction, and is inhibited so soon as it attempts the other. To account for them, we must suppose that stimulations which act upon the central region of the nerve-cell serve to propagate the inhibitory processes (the negative molecular work) there in progress to the peripheral region; while, conversely, stimulations which act upon the peripheral region effect a diffusion of the excitatory processes (the positive molecular work) there released over the central region. The intrinsic probability of this hypothesis is vouched for by the well known fact that in all chemical processes, in which the state of equilibrium of complex molecules has once been disturbed, the disturbance is normally transmitted to other molecules. The explosion of the very smallest quantity of nitrogen chloride is enough to decompose many pounds of this substance, and a single blazing chip may set a whole forest on fire. There is, it is true, an apparent difficulty in the case before us: molecular processes of opposite character are distributed over one and the same mass, according to the direction from which the stimulation proceeds. We must, however, remember that these processes are constantly in progress, side by side, in both regions of the cell; and that, as the constant exchange of materials demands, there is a continuous and gradual transition from the one region to the other. We may again return to the illustration of the nerve-fibre under the action of the constant current. In the neighbourhood of the anode there is a preponderance of inhibitory, in that of the cathode a preponderance of excitatory molecular processes. But it may be demonstrated, by aid of test-stimuli of varying intensity, that there is increase at the anode not only of the inhibitory but also of the excitatory processes; while, on the other hand, as the strength of current is increased, the inhibitory process is propagated to the cathode and beyond (cf. pp. 97 f.). Similarly [p. 99] with the nerve-cell. Fig. 34 may illustrate, e.g., the behaviour of the cells of the dorsal and ventral cornua of the myel to incoming and outgoing fibres. M represents a cell of the ventral, S a cell of the dorsal cornu; c and c' are their central p and p' their peripheral regions. In the ventral half of the myel, stimulation can travel  only from m' to m; in the dorsal half, only from s to s': a stimulus proceeding from m, s' is inhibited in c, c'. A stimulation passing between S and M can travel only in the direction from S to M, and not contrariwise; for a stimulus operating at m will be arrested in c, and a stimulus applied at m' may be conducted as far as c', but cannot go farther. Finally, the reflex excitation proceeding from s must be inhibited by a stimulation acting at s', because the molecular motion of inhibition arising in c' tends to spread over the peripheral region, and thus destroys, in whole or part, the excitation there set up. The morphological facts put it beyond question that the part of the ganglion cell here designated the central region is the place of origin of the axis-cylinder or neurite, and that the peripheral region gives rise to the dendrites. The latter region belongs accordingly to the actual periphery of the ganglion-cell, though it may perhaps also extend some little distance into the central ground-reticulum.[42]
only from m' to m; in the dorsal half, only from s to s': a stimulus proceeding from m, s' is inhibited in c, c'. A stimulation passing between S and M can travel only in the direction from S to M, and not contrariwise; for a stimulus operating at m will be arrested in c, and a stimulus applied at m' may be conducted as far as c', but cannot go farther. Finally, the reflex excitation proceeding from s must be inhibited by a stimulation acting at s', because the molecular motion of inhibition arising in c' tends to spread over the peripheral region, and thus destroys, in whole or part, the excitation there set up. The morphological facts put it beyond question that the part of the ganglion cell here designated the central region is the place of origin of the axis-cylinder or neurite, and that the peripheral region gives rise to the dendrites. The latter region belongs accordingly to the actual periphery of the ganglion-cell, though it may perhaps also extend some little distance into the central ground-reticulum.[42]
The results which follow from stimulation of peripheral ganglia, such as those of heart, blood-vessels, intestine, lend themselves readily to the same interpretation. Whether stimulation of the nerves which run to these ganglia produces excitation or inhibition depends likewise upon their mode of connexion with the nerve-cells. Thus, the inhibitory fibres of the heart will terminate in the central the accelerating fibres in the peripheral region of the ganglion-cells of this organ; it is not necessary to assume the existence of separate apparatus for the two processes. The result of stimulation can be modified in only one way. The ganglia are, at the same time that they are stimulated from without, in a state of continuous automatic stimulation, so that the incoming nerves can do no more than regulate the movements made. For the rest, the nerve-cells, here as before, show the phenomena of accumulation and summation of stimuli. Intensive excitation of the inhibitory nerves of the heart will, it is true, [p. 100] arrest the heart-beat after a very short interval; but stimulations of more moderate intensity can produce this effect only after several beats have been executed. The phenomenon appears still more plainly in the case of the accelerating nerves, where several seconds regularly elapse after the beginning of stimulation before acceleration sets in. On the other hand, the after-effect of the stimulus always persists for a considerable time after the stimulus itself has ceased to act; the heart returns only gradually to its original frequency of beat. There is also a further point in which the conditions here evidently differ somewhat from those obtaining in the skeletal muscles. In all regions of innervation that do not stand under direct voluntary control, the mechanisms which subserve the production and accumulation of excitatory and inhibitory effects are to be found, in part, in the muscles themselves; so that, under these circumstances, the muscle-substance is endowed, within limits, with the attributes elsewhere reserved for the peripheral and central nerve-substance.[43] In view of the close relation subsisting in other respects between nervous and contractile substance, we may perhaps look upon this result as a simple enhancement of the powers possessed by muscular tissue in its own right, an enhancement due to the independence attained by the peripheral organs.
We may now turn to the elementary phenomenon of practice which, as will be remembered, is exhibited by the peripheral nerve-substance. The phenomenon recurs, with modifications conditioned upon the law of propagation of molecular processes within the ganglion-cell, in the central substance. Its effects are by no means simple, as may be observed in the following instances. We find, on the one hand, that co-ordinated movements; whose first performance was difficult and required continuous voluntary control, gradually become easier and, at last, altogether in voluntary. We find, again, that functional disturbances, set up by the destruction of central elements, are gradually compensated, without restitution of these elements themselves. In the first of these phenomena, we have an increasing facilitation of the excitatory processes in consequence of their frequent repetition. The second suggests that, under suitable conditions, the stimulation may strike out new paths within the central substance: we may accordingly designate this latter effect of practice, in contradistinction to direct practice by repetition of function, as path-making or canalisation.[44] We are thus led to the following conclusions. First, when an excitatory process is frequently conducted through a ganglion [p. 101] cell in a given direction, the cell thereby acquires a prepotent disposition to conduct any future stimulations that may reach it in the same direction. Secondly, the processes of conduction in the central substance at large cannot be confined within fixed limits; elements in which, under normal circumstances, the excitations are annulled by concurrent inhibitions must be able, under the new conditions of practice introduced by the destruction of former conduction paths, to enter into new functional connexions. Translated into terms of the hypothesis developed above, this would mean that the frequent repetition of conduction, in a certain direction, so modifies the portion of the central substance which lies along this particular path that it takes on, more and more completely, the character normally attaching to the peripheral region. But, as a matter of fact, this sort of transformation is just what might have been predicted from the general laws of stimulation. We have seen that, in the peripheral nerve, the inhibitory forces are further and further reduced, under the action of repeated stimuli; so that at first, before functional capacity is exhausted, frequent repetition of stimulation means enhancement of irritability. Repetition of stimulus, that is, always and everywhere brings with it an alteration of the nerve-substance, which thereby loses the power of exerting the inhibitory influence connected with restoration of its internal forces. It is to this fact that we must turn for explanation of the principle of practice, in its special significance for the central functions, noting at the same time that the principle divides into two less general principles, each of importance for the understanding of these functions, and each in various ways supplementing the other: the principles of localisation and of vicarious function. We shall find, when we come to consider the functions of the central organs of the nervous system, that both alike are indispensable aids to an interpretation of the phenomena.
(b) -- Relation of Nervous to Psychical Processes
These considerations yield, however, another and a more general result important not only for the physiological but also for the psychological aspect of vital phenomena. We have taken, as a measure of the effects which the nervous substance can produce within itself and can transmit to other elements of the body, resembling it in certain general properties, the effects exhibited by the muscle; and we have done this, partly because the muscular effects are most easily accessible to observation, partly because they can be subsumed, with the least possible ambiguity, to metric principles of universal validity. Now we have no right to suppose that the laws which govern the transference of nervous molecular processes to the contractile substance are at all different from the laws which regulate [p. 102] their transmission to other substances, whose properties show them to be related to the nervous elements, -- more especially, therefore, to the substances that are of peculiar import for the psychical aspect of vital phenomenon, the elements of the sense-organs. On the contrary, the identity of these laws is a matter of course. It follows, therefore, that the changes set up by the action of stimulus in the sensory cells, and in the peripheral and central portions of the nervous system connected with them -- whether the stimulus be applied from without or arise within the system itself -- consist always in those forms of positive and negative molecular work whose general laws we have sought to trace in the symptoms presented by the muscular system. We have seen that all these forms can readily be brought under the general point of view of the principle of energy; we have had, as illustrations of them, the decomposition and recomposition of chemical compounds, the liberation and absorption of heat, the increase and decrease of actual mechanical work. Now the processes thus analysed remain, always, physical and chemical processes. It is never possible to arrive, by way of a molecular mechanics, at any sort of psychical quality or process. If, then, experience teaches us that the molecular processes within our nervous system may have psychical concomitants, we can only say that we are here in presence of a fact which lies altogether beyond the cognisance of a molecular mechanics of nerve-substance, and consequently beyond the cognisance of any strictly physiological inquiry. It would fall within the scope of physiology only if we were able in some way to interpret the psychical processes themselves as molecular processes, i.e. in the last resort, as modes of motion or as physical energies. This, however, we cannot do: the attempt fails at once, under whatever guise it may be made. Psychical processes refuse to submit to any one of our physical measures of energy; and the physical molecular processes, so far as we are able to follow them, are seen to be transformed, variously enough, into one another, but never directly into psychical qualities. In saying this, we do not, of course, reject the idea that psychical processes may be regularly attended by an interchange of physical forces, which as such forms a proper object of co-ordinate investigation by the molecular mechanics of the nervous system; nor do we deny, what would naturally follow, that psychical symptoms may he taken as indicative of definite physiological molecular processes, and that these in their turn, if it ever happens that we know more about them, may be taken, under certain circumstances, as indicative of psychical conditions. But such a relation between the two departments is entirely compatible with their separate independence, with the impossibility, at any time or by any means, of the reduction of the one to the other. As a matter of fact, we can no more derive the mechanics of nerve-substance from the connexions and relations [p. 103] of our sensations and feelings, than we can derive the latter from molecular processes. We have, then, no choice as to the road which we shall take in the following Chapters. We must first of all occupy ourselves with the investigation of the bodily substrate of the mental life as a physiological problem; our task being, in the main, simply to apply the principles which we have discovered in the general mechanics of nervous substance to the complex connexions of nervous elements presented in the nervous system of the animals, and more especially of man. Psychological facts will here be accorded merely a symptomatic importance, in the sense defined above: we shall depart from this rule only when the critical discussion of certain hypotheses of psychological character, which have taken shape within nerve-physiology, requires us to raise the question whether and how far these hypotheses receive adequate support from the physiological facts themselves. But the general question, as to the nature of the relations which unite the mechanics of nerve-substance and of its complex effects in the nervous system, on the one hand, and the phenomena of the mental life, on the other, -- this question presupposes the analysis of both sets of facts, the physiological and the psychological: so that its investigation must, naturally, be postponed to the conclusion of the present work.
Notes
[1] A good summary of the most important facts regarding the mechanical properties of muscle will be found in TIGERSTEDT'S Lehrbuch der Physiologie, ii., 98, 128 ff. The reader should compare with this the recent papers of ROLLETT (PFLÜGER'S Arch f. d. ges. Physiol., lxiv. and lxxi.), SCHENCK (ibid. lxii., lxiii., lxiv., lxv., lxvii., lxxii.) and KAISER (Zeitschr. f. Biologie, xxxiii., xxxv., xxxvi., xxxviii.).
[2] CLAUSIUS, Abhandlungen zur mechanischen Wärmetheorie, ii., 1867, 214.
[3] Muscle-curves of this kind are termed, as proposed by A. FICK, 'isotonic' (curves of equal tension), and distinguished from the 'isometric' curves, described when the muscle is prevented, by over-loading, from making contractions of any considerable extent (cf. Fig. 26, p. 70). There is, of course, no such thing as a purely isotonic or purely isometric curve. In the twitch of the muscle, there must necessarily occur changes of tension, increasing in general with the amount of load, while an absolutely isometric muscle would not describe any curve whatsoever. Curves may also be obtained under the further condition that the tension increases during contraction, as when the muscle is made to pull against a spring ('auxotonic' curve), or that it is suddenly augmented during contraction by the application of a load, and so on. The different properties of the resulting curves are, however, of interest only for a mechanics of muscle.
[4] Cf. my Untersuchungen zur Mechanik der Nerven und Nervencentren, Abth. i., 1871, 177. The increase of the height of the muscle-curve with the distance of the point of stimulation from the muscle, first observed by PFLÜGER (Untersuchungen über die Physiologie des Elektrotonus, 140), has peen referred by many physiologists, following HEIDENHAIN (Studien des physiol. Instituts zu Breslau, i., 1), to the effect of the section or, where connexion with the myel is retained to the unequal decay of the nerve-tissue. If this hypothesis be correct, we must suppose that the excitability of the living nerve is the same at all points along its course. I have, however, shown, and the observation has been subsequently confirmed by TIEGEL (PFLÜGER'S Arch. f. d. ges. Physiol., xiii., 598), that the greater excitability of the parts more remote from the muscle obtains also in a living animal in which the circulation is maintained. I found, in particular, that the lengthening of contraction, which I had myself observed to be connected with increased length of nerve, is especially noticeable in the living nerve. This is, no doubt, the reason that it was not seen by experimenters who worked only with muscle-nerve preparations.
[5] Experiments on the superposition of two contractions were first made by HELMHOLTZ (Monatsber. d. Berliner Akad., 1854, 328). He found in opposition to the results noted above, that there was never anything more than a simple addition of the contractions. The greater heightening of the curve of summation has, however, been confirmed by KRONECKER and STANLEY HALL (Archiv. f. Physiologie, 1879, Supplementband, 19f.). The later experiments of M. VON FREY (ibid. 1888, 213) and J. VON KRIES (same vol. 537) also agree in all essential points with my own results.
[6] Mechanik der Nerven, i., 109 ff.
[7] Mechanik der Nerven, i., 63, 100.
[8] Mechanik der Nerven, i., 86, 190, 200.
[9] WUNDT, Arch. f. Anatomie u. Physiologie, 1859, 537; 1861, 781. Untersuchungen zur Mechanik der Nerven, i., 177 ff.
[10] HELMHOLTZ, Monatsber. d. Berliner Akademie, 1854, 328. These phenomena of summation are discussed with more accuracy of detail as regards the time-relations of the component stimuli by J. VON KRIES, in DU BOIS-REYMOND'S Arch. f. Physiologie, 1888, 538.
[11] WUNDT, Arch. f. Anatomie u. Physiologie, 1859, 549.
[12] BOWDITCH, Journal of Physiology, vi., 1887, 133. Arch. f. Physiologie, 1890, 505.
[13] PLÜGER, Untersuchungen über die Physiologie des Elektrotonus, 1859. WUNDT, Untersuchungen zur Mechanik der Nerven, i., 223 ff.
[14] A. FICK, Mechanische Arbeit und Wärmeentwickelung bei der Muskelthätigkeit, 1882.
[15] BERNSTEIN, Monatsber. d. Berliner Akademie, 1880, 186.
[16] According to BERNSTEIN'S investigations, PFLÜGER'S Arch. f. d. ges. Physiol., i., 190; Untersuchungen über den Erregungsvorgang im Nerven- und Muskelsysteme, 1871, 30.
[17] The negative variation of the muscle-current is of somewhat longer duration; it lasts approximately 0.004 sec. (BERNSTEIN, Untersuchungen, 64.) This time however, also falls within the limits of the period of inexcitability.
[18] BIEDERMANN, Elektrophysiologie, 1895, 71 ff.
[19] It is therefore advisable, in order to induce a reflex excitability that shall suffice for experimental series of some length, to help things out by minimal doses (0.002 to at most 0.004 mg.) of strychnine. I have convinced myself, by experiments specially directed upon the point, that minimal quantities of this poison do not affect the temporal course of the reflex contractions. See Untersuchungen zur Mechanik der Nerven und Nervene ntnen, ii., 1876, 9.
[20] Exceptions to this rule may occur, though very rarely, in cases of minimal reflex excitation and minimal motor stimulation: op. cit., 21.
[21] Op. cit., 14, 30, 37.
[22] Op. cit., 45 f.
[23] Cf. Note 1, p. 86, above.
[24] KRONECKER and STIRLING, Berichte d. k. sächs. Ges. d. Wissensch. zu Leipzig, math.-phys. Cl., 1874, 372. These observers declare, further, that the reflex twitch is invariably distinguished from the simple muscle-contraction by its more tetanic character (Arch. f. Physiologie, 1878, 23). I cannot follow them in this statement. It may evidently be explained by the fact that KRONECKER and STIRLING did not avail themselves of the minimal doses of poison referred to above, and were therefore obliged to use stronger stimuli for the excitation of reflexes. I would not, however, be understood to maintain that any hard and fast line can be drawn between simple contraction and tetanus at large. On the contrary, the acceleration of the course of the simple muscle-contraction, in its ascending branch, proves that even in it several successive excitatory impulses are at work.
[25] BEAUNIS, Rech. expér. sur les conditions de l'activité cérébrale et sur la physiologie des nerfs, 1884, 106.
[26] HELMHOLTZ, Monatsberichte d. Berliner Akademie, 1864, 307.
[27] KRONECKER and STANLEY HALL, Arch. f. Physiologie, 1879. Supplementband, 12. Similar observations are recorded by HORSLEY and SCHAEFER (Journ. Of Physiology, vii. 96), and, on the human subject, by GRIFFITH (ibid., ix. 39).
[28] HELMHOLTZ and BAXT, Monatsber. d. Berliner Akad., 1867, 228; 1870, 184 Experiments by VON KRIES (Arch. f. Physiol., 1886, Supplementband. 1 ff.) gave like results.
[29] Untersuchungen zur Mechanik der Nerven, ii., 64.
[30] Op. cit. 56 f. ROSENTHAL (Monatsber. d. Berliner Akad., 1873, 104; 1875, 419) speaks of a decrease of the latent period in strychnine tetanus, and BIEDERMANN (Elektrophysiologie, 1895, 501) accepts his statement. I do not understand this result; though with a high degree of strychnine poisoning, and with stimuli of moderate intensity, the increase of the latent period is not so pronounced as to be obvious at once, without the aid of some chronometric instrument.
[31] SETSCHENOW, Physiol. Studien über die Hemmungsmechanismen für die Reflexthätigkeit des Rückenmarks, 1863. SETSCHENOW and PASCHUTIN, Neue Versuche am Hirn und Rückenmark des Frosches, 1865.
[32] HERZEN, Sur les centres modérateurs de l'action reflexe, 1864, 32. SETSCHENOW, Ueber die elektrische und chemische Reizungder sensibeln Rückenmarksnerven, 1868, 40.
[33] GOLTZ, Beiträge zur Lehre von den Functionen der Nervencentren des Frosches, 1869, 44, 50. That other parts of the brain, besides those designated by SETSCHENOW, are able to inhibit reflexes, was demonstrated by GOLTZ in his croak experiment. Frogs whose cerebral hemispheres have been removed may be made to croak, with almost mechanical certainty, by a gentle stroking of the skin of the back; while with uninjured animals the same procedure very frequently fails of its effect. It appears, then, that the cerebral hemispheres also have the power of inhibiting reflexes (GOLTZ, Op. cit., 41). Experiments made by LANGENDORFF (Arch. f. Physiol., 1877, 133) and BÖTTICHER (Ueber Reflexhemmung, in Sammlung physiol. Abhandl., ii. Reihe, Heft 3) show that the same result may be obtained by blinding the animals.
[34] Untersuchungen, etc., ii., 84 ff., 106 ff. Morphine, on the other hand, seems, at a certain stage of its action, to increase the central inhibitions. For it was found by HEIDENHAIN and BUBNOFF that the contractions produced in animals by stimulation of the motor areas of the cerebral cortex were, in the normal state, enhanced, but in morphine narcosis inhibited, by mechanical stimulation of the skin. See PLÜGER'S Arch. f. d. ges. Physiol., xxvi., 137 ff.
[35] E. CYON has turned this idea to account for a theory of the central inhibitions: Bulletin de l'Acad. de St. Pétersbourg, vii., Decr., 1870. The facts which he adduces in support of it, so far as they are taken from the phenomena of vascular innervation, have been called in question by HEIDENHAIN. PFLÜGER'S Arch. f. d. ges. Physiol., iv. 551.
[36] BRONDGEEST, Over den Tonus der willekeurigen Spieren, Utrecht, 1860.
[37] CYON, Berichte d. k. sächs, Ges. d. Wiss., math.-phys. Cl., 1865, 86. Cf. on the other side, G. HEIDENHAIN, in PFLÜGER'S Arch. f. d. ges. Physiol., iv., 1871, 435.
[38] H. E. HERING and SHERRINGTON, in PFLÜGER'S Arch. f. d. ges. Physiol., lxviii., 1897, 222 ff.
[39] Cf. p. 53, above.
[40] Cf. pp. 55 f., above.
[41] Cf. the discussions in Part V. of the abnormal enhancement of excitability in the cerebral cortex, which presumably underlies certain forms of derangement of consciousness (dream, hypnosis).
[42] See above, Ch. ii, pp. 42 ff. I may be permitted to remark that this theory of the directions of central conduction as formulated, on the ground of purely physiological considerations (Untersuchungen, etc., ii., 1876, 116), long before RAMON Y CAJAL used the morphological facts to develope his views of the functional significance of the twofold origin of the nerve-fibres. Cf. also p. 50.
[43] T. W. ENGELMANN, PFLÜGER'S Arch. f. d. ges. Physiol., lvi., 1894, 149 ff.
[44] I take this very useful term Bahnung from S. EXNER, who first proposed it (Entwurf einer physiologischen Erklärung der psychischen Erscheinungen, i., 1894, 76), without meaning thereby to commit myself in any way to the views and hypotheses put forward by this author. (As the word 'facilitation' does not fit the present passage, we seem to have no better English term than 'canalisation.' -- TRANSLATOR.)