Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3173
(Return to index)
Principles of Physiological Psychology
By Wilhelm Wundt (1902)
Translated by Edward Bradford Titchener (1904)
CHAPTER II
Structural Elements of the Nervous System
§1. Morphological Elements
THE nervous system is made up of three kinds of morphological elements: (1) cells of peculiar form and structure, the nerve-cells or ganglion cells; (2) fibrous structures, originating as outgrowths from the cells, -- the nerve-fibres; and (3) a ground-reticulum, which in places is finely granular and in places fibrillar, and which consists of the terminal ramifications of the nerve-fibres and processes of the nerve-cells. To these must be added (4) a sustentacular substance, fibrous or amorphous in structure, which is regarded as a form of connective tissue.[1] The nerve-cells, with the fibrillar ground-reticulum that surrounds them, are essential constituents of all the central parts. In the higher nervous centres, however, they are restricted to definite areas, which, partly from their rich supply of capillary blood-vessels and partly from the presence of pigment-granules, collected both in the protoplasm of the cell-bodies and in the ground-reticulum, possess a darker coloration than the surrounding tissue. This grey substance contrasts so sharply with the white or myelinic substance that the distribution of cell-groups through the central organs may readily be followed by the naked eye. The myelinic substance itself owes its peculiar character mainly to the myelinic sheaths which enclose the nerve-fibres issuing from the grey substance. The connective tissue cement-substance occurs in three principal forms. As a soft and for the most part amorphous mass, the neuroglia, it serves to support the central nerves and cells. In the form of endoneurium and perineurium,[2] a denser tissue, showing tendon-like fibrillation, it extends among and surrounds the peripheral nerves. As the primitive sheath of Schwann, a membrane of glassy transparency and great elasticity, nucleated at intervals, it encases nearly all peripheral and a portion of the central nerve-fibres. These cement-substances form a sustentacular framework for the nervous elements. They serve, further, to carry the blood-vessels. And the perineurium [2][p. 40] imparts to the peripheral nerves, which have no solid wall of bone to protect them, the necessary power of resistance to mechanical injury.
(a) -- The Nerve-Cells
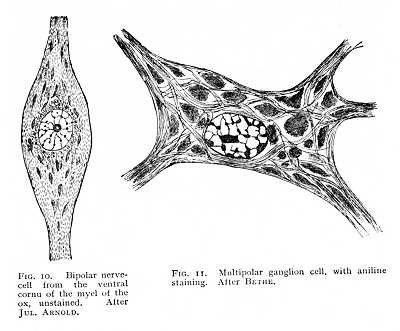 It is probable that the nerve-cells (Figg. 10-14) are everywhere devoid of a true cell-cortex. They vary in form from spherical to irregularly angular, and differ so extraordinarily in size that some can hardly be distinguished with certainty from the minute
corpuscles of the connective tissue, while others are visible to the naked eye. A clear nucleus, plainly vesicular in form, and provided with a large nucleolus, stands out in sharp contrast to the dully pigmented protoplasm. In the central organs the cells are embedded directly in the soft substance of the supporting tissue; in the ganglia, they are usually surrounded with an elastic sheath of connective tissue, often directly continuous with the primitive sheath of a nerve-fibre proceeding from them. The nerve-cells are characterised by their processes, one of which usually passes over directly into a nerve fibre, while the others ramify, if not immediately, after running a brief course, into fine fibrils. The former is called the axis-cylinder, nerve-[p. 41] process or neurite; the latter are termed protoplasmic processes or dendrites. Secondary dendritic processes may also arise, not from the cell itself, but from its neurite (Fig. 14, c). They are then named collaterals. The two types of process are shown with special clearness in many of the larger cells of the myel (spinal cord) and brain of vertebrates.
It is probable that the nerve-cells (Figg. 10-14) are everywhere devoid of a true cell-cortex. They vary in form from spherical to irregularly angular, and differ so extraordinarily in size that some can hardly be distinguished with certainty from the minute
corpuscles of the connective tissue, while others are visible to the naked eye. A clear nucleus, plainly vesicular in form, and provided with a large nucleolus, stands out in sharp contrast to the dully pigmented protoplasm. In the central organs the cells are embedded directly in the soft substance of the supporting tissue; in the ganglia, they are usually surrounded with an elastic sheath of connective tissue, often directly continuous with the primitive sheath of a nerve-fibre proceeding from them. The nerve-cells are characterised by their processes, one of which usually passes over directly into a nerve fibre, while the others ramify, if not immediately, after running a brief course, into fine fibrils. The former is called the axis-cylinder, nerve-[p. 41] process or neurite; the latter are termed protoplasmic processes or dendrites. Secondary dendritic processes may also arise, not from the cell itself, but from its neurite (Fig. 14, c). They are then named collaterals. The two types of process are shown with special clearness in many of the larger cells of the myel (spinal cord) and brain of vertebrates.
The nerve-fibres do not form independent elements of the nervous system. They originate, as embryology teaches us (Fig. 9), in outgrowths from nerve-cells, and they remain throughout in connexion with the cells whose processes they are. We may accordingly consider the nervous system in its entirety as a vast conglomerate of nerve-cells, all woven together by fibrillar runners. Under these conditions, the only processes of the central cells that attain to any measure of apparent independence, as fibrillar elements, are those entering into connexion with the peripheral organs. But even the fibrils of the muscular and cutaneous nerves, which in many cases extend without break over large distances, are really nothing more than cell processes long drawn out. It is, therefore, the nerve-cell that is the main variable in the nervous system. Both in number and nature of its processes and in its own internal structure, the cell evinces characteristic differences, often strongly marked, from one part of the nervous system to another.[3] When highly magnified, most nerve-cells show, even without treatment by selective reagents, a fibrillated structure; clusters of granules are set, in scattered masses, between the meshes of this fibrillar network, and a special network of granules and fibrillae encloses the nucleus (Fig. 10). The granular deposits are named, from their discoverer, the corpuscles of Nissl; they are also known as tigroid bodies, or as chromophilous substance. Colour-staining brings them out with greater clearness, since they have an affinity for the dyes of the histologist, while the fibrillae and the amorphous ground-substance remain unaffected (Fig. 11). It appears, further, that these bodies stand in a peculiar relation to the different forms of cell-process; they are assembled in greater numbers at the points of origin of the dendrites, but are entirely absent from the part of the cell that gives off the neurite or axis-cylinder (Fig. 12, lower right-hand portion). Finally, besides this network of fibrillae which run their course within the substance of the cell, and whose continuity with the cell-processes evidences their nervous character, there is sometimes found a pericellular reticulum, which, basket-like, encloses the whole outer wall of the cell. Its fibrillae can, in most cases, be traced [p. 42] into the dendrites, so that they too are, in all probability, to be looked upon as nervous structures (Fig. 13).
Nerve-cells are classified, according to the number of processes they send out, as unipolar, bipolar and multipolar. 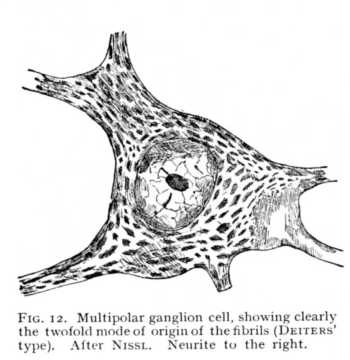
Unipolar cells are, however, always of rare occurrence; and, where they occur, have probably arisen secondarily, in course of growth from the originally bipolar form, by a fusion of its two processes, -- which, we may note, divide again immediately after their emergence from the cell (see Fig. 21, z, p. 50). The bipolar cell is found more especially in the peripheral regions, e.g. in the spinal ganglia, in the retina, and (to some extent) in the ganglia of the sympathetic system. The great majority of nerve-cells are, however, multipolar. As a rule, every such cell gives off a single neurite, and an indeterminate number of dendrites. The divergent characters not only of the processes themselves, but also of the portions of the cell with which they are connected (Fig. 12) render it, in the present case, an exceedingly probable hypothesis, that the difference of structure is paralleled by a corresponding difference of function. As a matter of fact; the fibrils of the large cells of the ventral cornua of the myel that pass over into the motor nerves, are without exception neuritic; while the processes that tend from the same cells towards the higher regions of the myel are dendritic in nature. RAMON Y CAJAL has accordingly suggested that the dendrites are devoted exclusively to cellipetal, the neurites to cellifugal conduction.[4] This scheme can, however, hardly he applied to all nerve-cells, without exception, since there are many cases in which no clear difference between the various cell-processes can be made out.
For the rest, over and above their different manner of origination from the cell body, their shorter course, and their greater wealth of branches, the dendrites are morphologically distinguishable from the neurites by their character as 'protoplasmic' processes; their irregular [p. 43] nodosity (Fig. 14) suggests the pseudopodial processes of the Rhizopoda (Fig. 2). They have also been observed, under the action of mechanical, chemical or electrical stimulation, to make amoeboid movements; though it 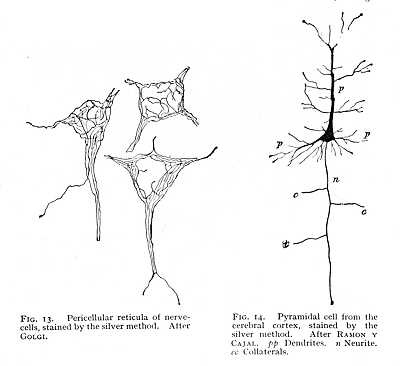 is doubtful whether these changes are to be interpreted as vital phenomena, on the analogy of the contraction of protoplasm and of muscular tissue, or whether they are not rather simply the direct physical and chemical effects of the stimuli applied.[5]
is doubtful whether these changes are to be interpreted as vital phenomena, on the analogy of the contraction of protoplasm and of muscular tissue, or whether they are not rather simply the direct physical and chemical effects of the stimuli applied.[5]
These differences between the two kinds of cell-processes are, however, as we said above, not equally well marked in all cases. In particular, the difference in length and character of course may be comparatively slight, or may even disappear altogether, the neurite, like the dendrite, dividing after a brief period into a large number of delicate branches. It is also not uncommon to find cells, especially cells of small size, whose processes show no distinct sign of difference, of whatever sort. The cells [p. 44] with processes of markedly different form are usually termed, from their discoverer, the cells of DEITERS (Fig. 12); cells with quickly dividing neurites are known as cells of GOLGI'S type; and the cells without marked distinction of the processes are called intermediary or intercalary cells.[6]
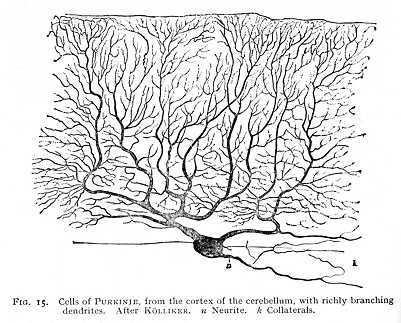
Finally, the dendrites, like the neurite, evince certain structural differences. Sometimes, as in the pyramidal cells of the cerebral cortex (Fig. 14) they divide without much complication, their branches tending in definite directions. Sometimes, again, as in the large PURKINJE cells of the cerebellar cortex (Fig. 15), their ramifications are exceedingly complex and widely extended.
(b) -- The Nerve-Fibres
We have seen that the nerve-process issuing from the nerve-cell forms the basis of the nerve-fibre. The main differences in the structure of the nerve-fibres depend upon differences in the character of the investing [p. 45] substances, which envelope the original neurite as it proceeds on its way. The constant constituent of a nerve-fibre, as follows at once from its mode of origin, is the neurite or axis-cylinder that forms the direct continuation of the nerve-process of a cell. The neurite is enclosed, first of all in the myelinic sheath, a substance which after death breaks up by a process of decomposition into bulbous masses; later in its course, it becomes surrounded by a structureless membrane, supplied at intervals with nuclei, -- the primitive sheath of SCHWANN (Fig. 17). Most of the central nerve-fibres possess a myelinic sheath, but no primitive sheath; and within the grey 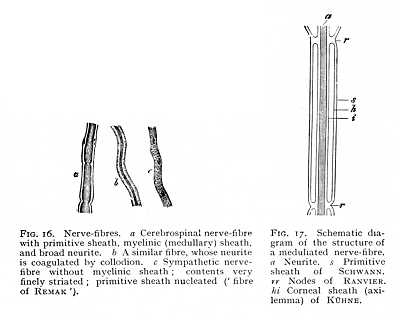 substance the myelinic sheath itself not seldom disappears. In the sympathetic system, on the other hand, the neurite is, as a rule, enveloped directly by the nucleated primitive sheath, and lacks the intervening myelinic substance (Fig. 16, c). (With few exceptions,) the nerve-fibres of invertebrates evince this constitution throughout. Lastly, the terminal branches of the nerves in the peripheral end-organs often break up into arborisation, consisting simply of fine neuritic fibrils.
substance the myelinic sheath itself not seldom disappears. In the sympathetic system, on the other hand, the neurite is, as a rule, enveloped directly by the nucleated primitive sheath, and lacks the intervening myelinic substance (Fig. 16, c). (With few exceptions,) the nerve-fibres of invertebrates evince this constitution throughout. Lastly, the terminal branches of the nerves in the peripheral end-organs often break up into arborisation, consisting simply of fine neuritic fibrils.
The two innermost of the three principal constituents of the nerve-fibre, myelinic sheath and neurite, possess a composite structure. If we trace a fibre throughout any considerable portion of its extent, we find that the myelinic substance does not afford a continuous investment of the neurite. The primitive sheath undergoes constriction at more or less regularly recurring intervals (nodes of RANVIER), and the [p. 46] myelinic sheath is thus divided up into cylindrical sections, separated by transverse partitions (Fig. 17). Since each section carries but a single cell-nucleus, we may suppose that it represents one of the cells of which the sheath is ultimately composed (Fig, 12). Within this internodal space (bounded by r r in the Fig.) there is, further, according to some observers, another double sheath, composed of a substance akin to epithelial tissue, and separating the neuritic thread from the myelinic sheath (hi).[7] While the myelinic sheath is thus subdivided, the neurite itself runs uninterruptedly front its point of origin to the conclusion of its course. It is made up, as was first observed by MAX SCHULTZE, of numerous primitive fibrils, which in many places, and especially where it issues from the nerve-cell, give it a finely striated appearance.[8] It is probable that these primitive fibrils pass, in the peripheral nerve terminations, into the dendritic arborisation into which many nerve-fibres are ultimately resolved.
Putting all this together, we may infer that the neuritic thread is the constituent of the nerve-fibre essential to the conduction of nervous processes; that the myelinic sheath discharges not a nervous but a nutritive function; and that the remaining investments are merely protecting structures.[9] The inference is borne out by the fact that the formation of the myelinic sheath follows at a comparatively long interval, in the development of the nervous system, upon the appearance of the neuritic thread. At the same time, there can be no doubt of its great importance. The fibres that are to become myelinic give no clear indication of irritability, or of functional capacity at large, until myelinisation is complete.[10]
The nerve-processes and the nerve-fibres that proceed from them are, then, extremely important for the connexion of the nerve-cells with the peripheral appendages of the nervous system, the sense-organs, glands, muscles, etc. But they never mediate a direct connexion between cell and cell. Wherever such connexion occurs, it appears to be mediated solely by the contact into which dendrites and collaterals are brought with one another throughout the grey substance. This view finds support in observations made upon the peripheral terminations of the nerve-fibres.[p. 47]
(c) -- Peripheral Nerve Terminations
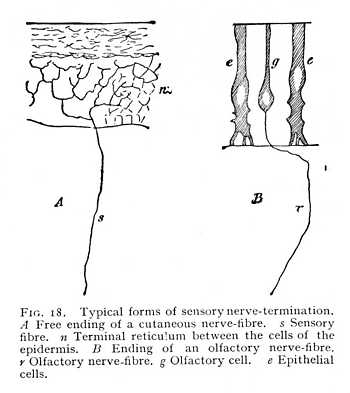 The termination of a nerve in the peripheral organs may take one of two forms. Either the ends of the neuritic threads divide up into a fascicle or network of finest dendritic fibrils, that terminate freely along the elements of other, non-nervous tissues; or the neuritic thread passes directly over into a terminal cell situated within or between the organs. The terminal cell may be an
original nerve-cell pushed out towards the periphery of the body; or it may have acquired this character later on in the course of development, by the penetration of a nerve fibril into an epithelial cell. The two forms of nerve-termination occur side by side, in these their characteristic differences, in the different sense-organs, where they are evidently connected with essential differences in the mode of sensory excitation. The first form shows most plainly in the terminations of sensory nerves in the skin. The neurite, as soon as it enters the lowermost epithelial layer of the cutis, breaks up, into a reticulum of delicate fibrils, whose dendritic branches surround the separate epithelial cells (Fig. 18, A). In some cases, it is true, this arrangement is so modified as to approximate more or less closely to the second form: there are cutaneous nerve-fibres whose fibrils penetrate the cells of the epidermis, or pass into or between the cells of the deeper lying connective tissue, and thus transform these originally non-nervous elements into peculiar sense-organs (touch-cells, end bulbs, touch-corpuscles, etc.). The nerve-terminations in the organ of hearing also follow, in the main, this cutaneous type.
The termination of a nerve in the peripheral organs may take one of two forms. Either the ends of the neuritic threads divide up into a fascicle or network of finest dendritic fibrils, that terminate freely along the elements of other, non-nervous tissues; or the neuritic thread passes directly over into a terminal cell situated within or between the organs. The terminal cell may be an
original nerve-cell pushed out towards the periphery of the body; or it may have acquired this character later on in the course of development, by the penetration of a nerve fibril into an epithelial cell. The two forms of nerve-termination occur side by side, in these their characteristic differences, in the different sense-organs, where they are evidently connected with essential differences in the mode of sensory excitation. The first form shows most plainly in the terminations of sensory nerves in the skin. The neurite, as soon as it enters the lowermost epithelial layer of the cutis, breaks up, into a reticulum of delicate fibrils, whose dendritic branches surround the separate epithelial cells (Fig. 18, A). In some cases, it is true, this arrangement is so modified as to approximate more or less closely to the second form: there are cutaneous nerve-fibres whose fibrils penetrate the cells of the epidermis, or pass into or between the cells of the deeper lying connective tissue, and thus transform these originally non-nervous elements into peculiar sense-organs (touch-cells, end bulbs, touch-corpuscles, etc.). The nerve-terminations in the organ of hearing also follow, in the main, this cutaneous type.
 The second form of nerve-termination is best illustrated from the organ of smell. Every olfactory nerve-fibre enters, in the olfactory mucous membrane, into a nerve-cell. This cell which lies between epithelial cells, [p. 48] is drawn out at its opposite pole, i.e. at the end turned towards the free sensory surface, into a thread-like continuation (Fig. 18, B). The nerve-terminations in the tongue and in the retina of the eye follow this second type. In both organs, the terminal fibrils are connected with sensory
cells. In their case, however, the sensory cells (taste-cells, retinal rods and cones) appear to be not true nerve-cells, but epithelial cells, which have been transformed into sense-cells by their connexion with nerve-fibres.[11]
The second form of nerve-termination is best illustrated from the organ of smell. Every olfactory nerve-fibre enters, in the olfactory mucous membrane, into a nerve-cell. This cell which lies between epithelial cells, [p. 48] is drawn out at its opposite pole, i.e. at the end turned towards the free sensory surface, into a thread-like continuation (Fig. 18, B). The nerve-terminations in the tongue and in the retina of the eye follow this second type. In both organs, the terminal fibrils are connected with sensory
cells. In their case, however, the sensory cells (taste-cells, retinal rods and cones) appear to be not true nerve-cells, but epithelial cells, which have been transformed into sense-cells by their connexion with nerve-fibres.[11]
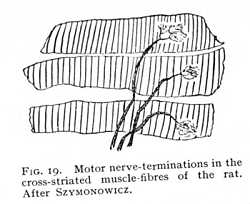
The nerve-endings in muscle conform in all essentials to the first of these types. Here too we observe, in the first place, a more or less elaborate division of the nerve-fibres that run to the separate elements of the muscular tissue.[12] In the muscles of reptiles, birds and mammals, the terminal fibrils finally branch out in a peculiar flattened prominence, the end-plate. Most observers place this structure within the transparent elastic sheath of the muscle-fibre, the sarcolemma, though some describe it as attached to the outer surface (Fig. 19).[13]
(d) -- The Neurone Theory
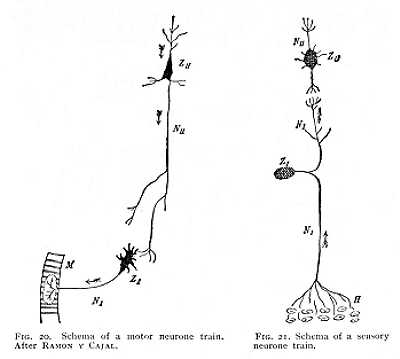
The facts which we have now passed in review as regards the nerve-cells, their processes, and the continuations of these processes into the peripheral organs appended to the nervous system, have led in recent years to the hypothesis that the conduction of nervous processes is mediated, in many cases, not -- as was formerly supposed -- by an unbroken continuity of the fibrillar elements, but rather by contact between the arborisations of the fibres of different nerve-cells. This hypothesis, it is needless to say, ascribes a greatly added importance to the nerve-cell. According to it, the functions of the nervous system are conditioned upon the spheres of function of the individual cells, -- the 'cell' in this sense including as an essential constituent the fibrillar elements issuing from the cell-body. We may therefore regard the nerve-cell together with its processes as the morphological, and presumably also as the functional unit, to which we [p. 49] are in the last resort referred for an understanding of the entire nervous system. This unit of nerve-cell with its dependent territory of fibrillar processes and arborisations, has been designated, on WALDEYER'S suggestion, a neurone. In the light of the neurone theory, the whole of the central nervous system, reaching with its appended organs to the extreme periphery of the body, appears as a system of such units, set side by side or arranged in ascending series: each unit maintaining a relative independence, from the unbroken continuity of its parts, and each connected with other similar units only contact-wise, by way of the terminal arborisations of the fibrils of the individual neurones.[14] Figg. 20 and 21 illustrate this conception, schematically, for two trains of neurones, a motor and a sensory, which may be taken as typical of the systems of conduction realised in the nervous system at large. The hypothetical scheme of the motor neurone train, given in Fig. 20, consists of two neurones, the one of which (NI), as motor cell (ZI) in the ventral cornu of the myel, is attached directly to a peripheral muscle-fibre (M), while the second (NII) belongs to a higher nervous centre. The neurite proceeding from the cell ZII gives off a certain number of collaterals, and finally resolves into fibrils that come into contact with the dendrites of the cell ZI. This cell in turn sends out a neurite, whose ramose fibrillar termination stands in contact with the motor end-plate of a cross-striated muscle-fibre. The hypothetical schema of the sensory neurone train, in Fig. 21, also shows two neurones: a peripheral, NI, that has its centre in a bipolar spinal-ganglion cell ZI, and a central neurone, NII, that belongs to a nerve-cell, ZII, lying somewhere in the higher regions of myel or brain. The neurone NI is connected by contact on the one side, through the terminal arborisation of its longer, peripherally directed fibre, with the cutaneous region H (cf. p. 47, Fig. 18 A), and on the other, through the dendrites of its second, upward trending process, with the neurone NII. These bimembral chains are, naturally, to be considered only as the very simplest schemata of neurone connexion. We must suppose in general that several neurones, now all lying at the same level and now arranged in ascending order, are united in the nervous centres to form neurone chains Where nerve-cells have been forced outward, as 'sensory cells,' into the peripheral organs, it is possible that there, too, similar arrangements may prevail. Indeed, as we shall see later on, the morphological conditions often point unequivocally to such peripheral neurone connexions (cf. below, Chs. V., VIII.).
Whether the individual cell territories are, always and everywhere, related to one another in the manner indicated by these diagrams is, we must [p. 50] admit, still an open question. So far, the neurone theory must be regarded simply as an hypothesis that brings together, in a very happy way, a large number of the data of current histology. Whether the definition of the neurone in general, and whether in particular the views of the interconnexion of the neurones promulgated especially by RAMON Y CAJAL, will prove to be tenable in all cases, cannot now be decided. Even at the present day, the theory does not want for opponents. Fortunately, the settlement of these controversies among the morphologists is not of decisive importance for a physiological understanding of nervous functions.
 Physiological interpretation must be based, first of all, upon the manifestations of function, and these can be brought, later on, into relation to the anatomical facts. The opposite plan, of erecting elaborate physiological -- not to say psychological -- hypotheses upon purely anatomical foundations, is, of course, to be rejected without further argument. From this point of view, however, it must be conceded that the idea of neurones, and the view that this idea [p. 51] suggests of a connexion between the central elements which is relatively variable, and in certain circumstances perhaps determinable by the exercise of the functions themselves, accords better with the facts than the older view of an uninterrupted continuity of the nerve-fibres, and its dogmatic corollary of isolated conduction, were able to do. We need appeal only to the observations on the possibility of vicarious functioning, and on the substitution of new conduction-paths for others that have for some reason become impracticable. The anatomical plan of neurone connexions is evidently more adequate than this older view to the physiological results which prove that there exists, along with a certain localisation of functions, a very considerable capacity for adaptation to changed conditions. More than this, -- more than an ex post facto representation of the course of events, -- the neurone theory, naturally, cannot give us. Should that theory fall, the acts of vicarious function and of new adaptation would still all remain as they were, and would still have to be brought somehow into agreement with the properties of the anatomical substrate of the functions involved.
Physiological interpretation must be based, first of all, upon the manifestations of function, and these can be brought, later on, into relation to the anatomical facts. The opposite plan, of erecting elaborate physiological -- not to say psychological -- hypotheses upon purely anatomical foundations, is, of course, to be rejected without further argument. From this point of view, however, it must be conceded that the idea of neurones, and the view that this idea [p. 51] suggests of a connexion between the central elements which is relatively variable, and in certain circumstances perhaps determinable by the exercise of the functions themselves, accords better with the facts than the older view of an uninterrupted continuity of the nerve-fibres, and its dogmatic corollary of isolated conduction, were able to do. We need appeal only to the observations on the possibility of vicarious functioning, and on the substitution of new conduction-paths for others that have for some reason become impracticable. The anatomical plan of neurone connexions is evidently more adequate than this older view to the physiological results which prove that there exists, along with a certain localisation of functions, a very considerable capacity for adaptation to changed conditions. More than this, -- more than an ex post facto representation of the course of events, -- the neurone theory, naturally, cannot give us. Should that theory fall, the acts of vicarious function and of new adaptation would still all remain as they were, and would still have to be brought somehow into agreement with the properties of the anatomical substrate of the functions involved.
The morphological differences between the processes of the nerve-cells, that have formed the point of departure for the development of the neurone theory, were first pointed out by DEITERS, in his work upon the large cells of the ventral cornua of the myel. GERLACH discovered the fibrillar structure of the intercellular substance, and HIS the embryological connexion of nerve-fibres with nerve-cells. GOLGI, KÖLLIKER, NANSEN, W. HIS, G. RETZIUS, RAMON Y CAJAL and many others have made the nerve-cell a subject of special investigation.[15]
It is but natural that the results obtained should not be always in agreement. GOLGI and NANSEN supposed that the dendrites are merely nutritive elements; and GOLGI held, further, that the interlacing fibres of the ground-reticulum anastomose to form a closed system. The other observers declared for the nervous character of the dendrites, and were unable to confirm the occurrence of anastomosis in the ground-reticulum. On the side of function, GOLGI propounded the hypothesis that the neurites pass exclusively into motor nerve-fibres, while the sensory nerves take their origin from the ground-reticulum. It would follow from this, since GOLGI did not recognise the nervous nature ot the dendrites, that the connexion between sensory and motor fibres is mediated not by any sort of nerve-cell, but only by the fibrillar substance of the ground-reticulum, and there, in all probability, by mere mechanical contact of the fibres. If on the other hand we admit, as the great majority of observers are now ready to do, that the dendrites are nervous in character, then we must suppose, as has been shown in particular by RAMON Y CAJAL, that while all centripetally conducting nerve-fibres first of all arborise into fibrils in the ground-reticulum, they afterwards avail themselves of the protoplasmic processes to discharge into nerve-cells. If this hypothesis be sound, the terms 'centripetal' and 'centrifugal' cannot be regarded as identical with 'sensory' and 'motor'; they are referable, in every case, only to the cells with which the fibres are con-[p. 52]nected. Centripetal, in this sense, are all conduction-paths that convey excitations to determinate nerve-cells; centrifugal, all conduction-paths that carry excitations from them. In general, therefore, the peripheral sensory nerves will belong to a centripetal, and the motor nerves to a centrifugal system. But within the central conduction-paths, i.e. those that run between different ganglionic systems, there may be fibres, centrifugal in respect of proximate cell-origin, that possibly possess a sensory character, and others, centripetal in origin, whose functions may possibly be motor. This view of the functions of the cell-processes evidently carries with it a relative independence of the territories of the individual nerve-cells, -- a phase of the subject to which WALDEYER especially has called attention, and which has led him to introduce the idea of the neurone. Most recent investigators adopt the neurone theory. At the same time, there has alas been a certain amount of dissent, based especially upon the oft repeated observation of the continuity of the fibrils within the nerve-cells.[16] It has even been maintained that the fibrils pursue an unbroken course throughout the entire nervous system, the nerve-cells included: an hypothesis first put forward by MAX SCHULTZE, the discoverer of fibrillar cell-structure,[17] and now revived on the ground of further work upon the same morphological phenomena.[18]
The structural schema, of RAMON Y CAJAL, and the neurone theory that is based upon it, stand in the forefront of recent neurological investigation. Anatomists have also devoted much attention to the finer structure of the nerve-cell itself. There have been two remarkable discoveries in this field, that have aroused especial interest: NISSL'S announcement of the tussock-like accumulations of granules (Figg. 10, 14),[19] and the observations made in many quarters on the fibrillar structure of the nerve-cells.[20] Neither of these, it is true, has passed unchallenged; both the granular masses and the fibrils have been explained as precipitates from the cell-substance, due to microchemical treatment or to post-mortem coagulation.[21] Nevertheless, the hypothesis that these structures exist in the living tissue is confirmed br the fact that they have been observed in fresh preparations, untreated by staining reagents (Fig. 10).[22]
NISSL'S corpuscles have further been observed to undergo noteworthy changes under the action of poisons, like arsenic, or as the effect of intense fatigue or other trophic disturbances. The tussocks decrease, both in size and in number, so that in many cases they can still be observed only at certain parts of the cell-body, while the nucleus becomes farther and farther dis-[p. 53]placed towards the cell-periphery, and finally disappears altogether. These changes correspond exactly to those observed in inflammatory conditions of the grey substance in the human brain, and termed homogeneous turgescence of the cells (Fig. 22 A). They suggest the idea that the tussocks discharge a specific function, intimately related to cell-nutrition. These structures are, perhaps, to be explained as accumulations of reserve material, to be drawn upon for functional purposes if this be true, we must probably attribute to them the trophic influence which the nerve-cell exercises upon the fibres proceedings from it, and which apparently makes the cell their nutritive as well as their functional centre.[23] This influence is shown by the fact that those fibres of a transsected nerve which remain connected with the central organ persist for a long time without change, whereas the fibres of the peripheral portion of the nerve, the part that is separated from the centre, very soon show signs of degeneration. First of 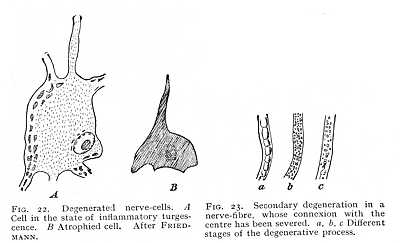 all, the myelinic contents of the fibre divides into clots (Fig. 23 a). Then, these clots, together with the neuritic fibrils, break up into granules (b). These in turn are slowly resorbed (c) until they altogether disappear: so that, finally, nothing is left of the nerve but its connective tissue investments.[24] It is, however, probable that the appearance of these degenerative processes is further hastened by the arrest of function which naturally follows from the sectioning of the nerve. This view is confirmed, on the one hand, by the fact that, after a very long time, the central end of the transsected nerve also becomes atrophied, and on the other by the observation that nerve-cells, which have been thrown out of function by sectioning of a nerve-trunk or by injury to the peripheral region supplied by them, gradually shrink up (Fig. 22 B). In the case of young animals especially, this cell-shrinkage sets in comparatively quickly, after extirpation of the region of nervous diffusion. It has also been observed in man, as a secondary atrophy of the nerve-centres.[25][p. 54]
all, the myelinic contents of the fibre divides into clots (Fig. 23 a). Then, these clots, together with the neuritic fibrils, break up into granules (b). These in turn are slowly resorbed (c) until they altogether disappear: so that, finally, nothing is left of the nerve but its connective tissue investments.[24] It is, however, probable that the appearance of these degenerative processes is further hastened by the arrest of function which naturally follows from the sectioning of the nerve. This view is confirmed, on the one hand, by the fact that, after a very long time, the central end of the transsected nerve also becomes atrophied, and on the other by the observation that nerve-cells, which have been thrown out of function by sectioning of a nerve-trunk or by injury to the peripheral region supplied by them, gradually shrink up (Fig. 22 B). In the case of young animals especially, this cell-shrinkage sets in comparatively quickly, after extirpation of the region of nervous diffusion. It has also been observed in man, as a secondary atrophy of the nerve-centres.[25][p. 54]
Such are the phenomena that occur as after-effects of enhancement or abolition of function in the nerve-cells and nerve-fibres. The changes observed as the results of stimulation in the dendritic processes, and interpreted by many observers as immediate manifestations of life, are of a very much more questionable nature. Amoeboid movements of the dendrites were first described by RABL-RÜCKHARD. They may possibly be explained as phenomena of imbibition and coagulation. At any rate, the psychophysical theories of sleep and waking, dissociation of consciousness, and what not, that certain authors have created upon them, are purely imaginary psychological constructions, based on an extremely scant and more than doubtful foundation of physiological observation.[26]
§2. Chemical Constituents
The chemical substances of which the morphological elements of the nervous system are composed are as yet but imperfectly known. The greater portion of the investing and sustentacular tissues -- the endoneurium and perineurium, the primitive sheath, and in part the neuroglia of the nerve-centres -- belong to the class of collogenic and elastic substances. The only exception is the corneal sheath surrounding the myelin, which is said to consist of a corneal substance allied to epithelial tissue, and termed neurokeratin.[27] The nerve-mass proper is a mixture of various substances, several of which resemble the fats in their solubilities, while they differ widely in chemical constitution. They have been found, not only in nerve-substance, but also in the corpuscles of blood and lymph, in egg-yolk, in sperma, and to a less degree in many other tissues. The most important of them is protagon, a highly complex body, to which LIEBREICH has assigned the empirical formula C116H241N4PO22. This formula is, naturally, intended merely to give an approximate idea of the extreme complexity of the chemical molecule of this compound.[28] From protagon are derived lecithin and cerebrin, decomposition-products which probably occur along side of it in the nerve-substance, and together with it form the myelin of the myelinic envelope. Lecithin, it is supposed, is not a single body of stable constitution, but consists of a series of compounds that resemble the compound ethers: substances which in physical and chemical con-[p. 55]stitution are closely allied to the fats, and in which the radicals of certain fatty acids, of phosphoric acid and of glycerin (a component of most of the animal fats) are combined with one another and with a strong amine base, cholin.[29] Lecithin has two characteristic properties. The large proportion of carbon and hydrogen which it contains gives it a high heat of combustion; and its complex nature renders it easily decomposable. Cerebrin, if boiled with acids, yields a sugar and other, unknown, decomposition products, and has accordingly been referred to the nitrogenous glucosides. Like lecithin, it is in all probability not a single body, but a mixture of several substances, which have been distinguished as cerebrin, homocerebrin and encephalin.[30] Lastly, cholesterin, a solid alcohol rich in carbon, which occurs in almost all the tissues and fluids of the body, plays a not inconsiderable part in the composition of nervous tissue. Besides these substances, which are all characterised by their high heat of combustion. Nervous tissue contains substances which are classed with the proteins, but of whose composition and chemical conduct very little is understood. Finally. it must be mentioned, as a characteristic difference between the grey substance of the nerve-centres and the white myelinic substance, that the former gives a weakly acid, the latter an alkaline or neutral reaction. The acid reaction appears, like that of the muscles, to be due to the presence of free lactic acid.[31] Some observers have, in fact, maintained that this free acid increases, as a result of activity, just as it does in muscle.[32] Apart from these differences of reaction, little is known of the distribution of the various constituents in the various elementary divisions of nervous tissue. Only so much is certain, that in the peripheral nerve-fibres the neurite has all the general characteristics of a proteid, while the myelinic sheath evinces those of the myelins. In the ganglion cells, too, the nucleus would seem, from its microchemical conduct, to consist of a complex albumin-like substance, while in the protoplasm there is a mixture of albuminoid materials with protagon and its associates. The same constituents appear, further, to penetrate in part into the intercellular reticulum.
These facts render it probable that nervous substance is the seat of a chemical synthesis, whereby the complex nutritive substances carried by the blood are ultimately transformed into compounds of still greater com-[p. 56]plexity, representing (as their high heats of combustion show) a very considerable amount of potential energy. This view of the chemism of nerve-substance is attested, first of all, by the appearance of protagon and the lecithins in such quantity that their production in situ is evidently far more probable than their deposition by the blood. The parent substances of protagon itself and of the bodies associated with it are to be sought, we must suppose, in the albumin-like substances of ganglion cell and neurite. There can, for that matter, be no doubt that the elementary structures of the animal body have the power of converting simpler proteids into more complex. Apart from the undisputed observation of synthetic processes within the body,[33] we have further evidence in the fact that substances containing phosphorus, which closely resemble the albuminates in their composition and chemical conduct, appear under conditions that definitely suggest their formation within the organic cell. A compound of this kind, nuclein, appears in particular to form the principal constituent of the cell-nuclei.[34] Hence we may say, tentatively, that the most important physiological result of the attempts so far made to penetrate the chemical constitution of the constituents of the nervous system is this and this only: that the chemism of nerve-substance is very particularly directed upon the formation of compounds possessing a higher heat of combustion or a larger store of potential energy, At the same time, the differences in the properties of the grey and white substance, scanty as they are, point to the conclusion that the central elements are the principal seat of the chemical processes which mediate the functions of the nervous system. These results, then, are practically all that we need bear in mind, as the outcome of chemical investigation of nervous substance up to the present time, when we approach the problems of the physiological mechanics of the nervous system.
Notes
[1] Connective tissue forms a part of the sustentacular tissue of the nervous system. But the neuroglia, which forms its larger part, is an ectodermic structure, with close relations to the neurogenetic tract. See G. A. PIERSOL, Normal Histology, 1893, 79. -- TRANSLATOR.
[2] The text has 'neurilemma' in both instances. This is now a synonym for the primitive sheath. -- TRANSLATOR.
[3] DEITERS, Untersuchungen über Gehirn u. Rückenmark des Menschen u. der Säugethiere, 1865, 53 f. HIS, Arch. f. Anatomie, Supplementband, 1890, 95 ff. VON LENHOSSEK, Der feinere Bau des Nervensystems, 2te Aufl. 1895, 36 ff. HELD, Arch. f. Anatomie, 1897, 204; Suppl. 273. BETHE, Arch. f. mikroskip. Anatomie, 1900, lv., 513. GOLGI, Verhandl. d. anatom. Gesellschaft auf d. 14 Vers. zu Pavia, 1900; Anat. Anzeiger, xviii., Ergänzungsheft.
[4] RAMON Y CAJAL, Les nouvelles idées sur la structure de systéme nerveux ches l'homme et chez les vertébrés, 1894.
[5] RABL-RÜCKHARD, Neurol. Centralblatt, 1890, 199. DUVAL, Soc. de Biologie, 1895. Cf. KÖLLIKER, Verh. d. Würzburger phys-med. Gesellschaft, 1895.
[6] These intermediate cells (intermediäre oder Schaltzellen) appear to correspond to what are sometimes termed the GOLGI cells of the first type, and the GOLGI cells of the text to the GOLGI cells of the second type. -- TRANSLATOR.
[7] EWALD and KÜHNE, Verhandl. d. naturhist.-med. Vereins zu Heidelberg, N. F. i. 5. The presence of this intermediate membrane in the living nerve-fibre is denied by T. W. ENGELMANN, in PFLÜGER'S Arch. f. d. ges. Physiol., xxii., 1880, 1 ff.; KÖLLIKER, Gewebelehre, 6te Aufl., ii., 13.
[8] MAX SCHULTZE, in STRICKER'S Gewebelehre, 1871, 108. POWER'S trs., i., 1870, 150.
[9] PIERSOL, Normal Histology. 63 f. -- TRANSLATOR.
[10] See below, Ch. v. § 2.
[11] For a detailed account of the central and peripheral terminations of the sensory nerves, see Chs. v. vii., below.
[12] PIERSOL, Normal Hisology, 90. -- TRANSLATOR.
[13] KÜHNE, in STRICKER'S Gewebelehre, ii., 1871, 682. POWER'S trs., i., 1870, 210. SZYMONOWICZ, Lehrbuch d. Histologie, 1901, 306. MACCALLUM'S trs., 1902, 312 ff.
[14] WALDEYER Ueber einige neuere Forschungen im Gebiet der Anatomie des Centralnervensystems, in the Deutsche med. Wochenschrift, 1891, nos. 44-48. For the history of the theory, see M. von LENHOSSEK, Der feinere Bau des Nervensystems, 2te Aufl., 1895, 103 ff.; M. VERWORN, Das Neuron in Anatomie und Physiologie, 1900.
[15] Cf. the bibliographies in M. VON LENHOSSEK, op. cit. 36 ff.; KÖLLIKER, Gewebelehre, 6te Aufl. ii. 5 ff.
[16] NISSL, Kritische Fragen der Nervenzellenanatomie, in the Biol. Centralblatt, 1896, 1898. HELD, Arch. f. Anatomie, 1897, 204; Suppl., 273. BETHE, Biol. Centralblatt, 1898, 10. 18. These authors believe, in general that the neurone theory affords an adequate idea of the earlier stages in the development of the nervous system; but that, at a later period, the processes of the individual cells oftentimes grow together, so that the original independence of the cell territories is not maintained.
[17] MAX SCHULTZE in STRICKER'S Gewebelehre, 1871, 108 ff. POWER'S trs., i. 172.
[18] APATHY, Biol. Centralblatt, 1889 and 1898 (vols, ix. and xviii.). Mittheilungen aus der Zool. Station zu Neapel, xii. 1897; also in Amer. Journ. of Insanity, lv., 1898, 51 ff.
[19] NISSL, Allg. Zeitschr. f. Psychiatrie, l., 1894.
[20] FLEMING, Arch. f. Mikrosk. Anatomie, xlvi., 1895, 373. LENHOSSEK, ibid., 345. MÖNCKEBERG and BETHE, ibid., liv., 1899, 135.
<{>[21] HELD, op. cit. Sometimes, as was discovered by BÜTSCHLI (Untersuchungen über mikroskopische Schäume und Protoplasma, 1892) and confirmed by HELD, a honey-combed appearance is presented both by the cell itself and by its nerve-process. HELD, however, regards this too as a result of coagulation.
[22] J. ARNOLD, Arch. f. mikrosk. Anatomie, lii., 1898, 542.
[23] NISSL, Allg. Zeitschr. f. Psychiatrie, xlviii., 1892. MARINESCO, Arch. f. Physiol., 1899, 89. VON WENDT, Skandin. Arch. f. Physio., xi., 1902, 372. M. FRIEDMANN, Neurolog. Centralblatt, 1891, 1.
[24] MÖNCKEBERG and BETHE, Arch. f. Mikrosk. Anatomie, liv., 1899, 135.
[25] GUDDEN, Arch. f. Psychiatrie, ii., 693.
[26] DUVAL, Hypothése sur la physiol. des centres nerveux, in the Comptes rendus de la société de biologie, 1895. SOUKHANOFF, La théorie des neurones, etc., in the Arch. de neurologie, 1897. QUERTON, Le sommeil hibernal et les modifications des neurones, Institut Solvay, Bruxelles, 1898.
[27] W. KÜHNE and CHITTENDEN, Zeitschr. f. Biologie, N. F. viii., 1890, 291.
[28] LIEBREICH, Ann. der Chemie u. Pharmacie, cxxxiv., 1865, 29. According to KESSEL and FREYTAG (Zeitschr. f. physiol. Chemie, xvii., 1893, 431), protagon further contains sulphur in its molecule. The views of these chemists, with the protagon theory at large, are sharply controverted by J. L. W. THUDICHUM (Die chemische Constitution des Gehirns des Menschen und der Thiere, 1901, 44 ff.). We cannot enter here into these differences of opinion. We can pass them over with the less scruple since they are, at present, without significance for the general relations of the chemism of nerve-substance to the physiological processes.
[29] The constitution of ordinary lecithin, according to DIAKONOW, is C44H90NPO9=distearyl-glycerin-phosphoric acid + trimethyl-oxethyl-ammonium-hydroxide. According to STRECKER, other lecithins may be formed in which the radical of stearic acid is replaced by some other fatty acid radical. See NEUMEISTER, Lehrbuch der physiol. Chemie, 2te Aufl., 1897, 91 ff.
[30] W. MÜLLER (Ann. d. Chem. u. Pharm. cv., 1858, 361) has worked oat for cerebrin the empirical formula, C37H33NO3. On the cerebrin series, cf. PARCUS, Journ. f. prakt. Chemie, 1881, 310; NEUMEISTER, Physiol. Chemie, 2te Aufl. 472.
[31] GSCHEIDLEN, in PFLÜGER'S Arch. f. d. ges. Physiol., viii., 1874, 71.
[32] MOLESCHOTT and BATTESTINI, Arch. de biologie ital., viii., 1887, 90.
[33] E. BAUMANN, Die synthetischen Processe im Thierkörper. Inaugural lecture. Berlin, 1878.
[34] MIESCHER, in HOPPE-SEYLER'S Physiologisch-chemische Untersuchungen, 4, 452; LUBAVIN, ibid., 463.