An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
CHAPTER IV
THE MENTAL LIFE OF THE MONKEYS; AN EXPERIMENTAL STUDY [1]
THE literary form of this monograph is not at all satisfactory to its author. Compelled by practical considerations to present the facts in a limited space, he has found it necessary to omit explanation, illustration and many rhetorical aids to clearness and emphasis. For the same reason detailed accounts of the administration of the experiments have not always been given. In many places theoretical matters are discussed with a curtness that savors of dogmatism. In general when a theoretical point has appeared justified by the evidence given, I have, to economize space, withheld further evidence.
There is, however, to some extent a real fitness in the lack of clearness, completeness and finish in the monograph. For the behavior of the monkeys, by virtue of their inconstant attention, decided variability of performance, and generally aimless, unforetellable conduct would be falsely represented in any clean-cut, unambiguous, emphatic exposition. The most striking testimony to the mental advance of the monkeys over the dogs and cats is given by the difficulty of making clear emphatic statements about them.
INTRODUCTION
The work to be described in this paper is a direct continuation of the work done by the author in 1897-1998 and described in Monograph Supplement No. 8 of the Psychological Review under the heading, 'Animal Intelligence; an Experimental Study of the Associative Processes in Animals.'[2] This monograph affords by far the best introduction to the present discussion, and I shall therefore assume an acquaintance with it on the part of my readers.
It will be remembered that evidence was there given that ordinary mammals, barring the primates, did not infer or compare, did not imitate in the sense of 'learning to do an act from seeing it done,' did not learn various simple acts from being put through them, showed no signs of having in connection with the bulk of their performances any mental images. Their method of learning seemed to be the gradual selection of certain acts in certain situations by reason of the satisfaction they brought. Quantitative estimates of this gradualness were given for a number of dogs and cats. Nothing has appeared since the 'Experimental Study' to negate any of these conclusions in the author's mind. The work of Kline and Small[3] on rodents shows the same general aspect of mammalian mentality.
Adult human beings who are not notably deficient in mental functions, at least all such as psychologists have observed, possess a large stock of images and memories. The sight of a chair, for example, may call up in their minds a picture of the person who usually sits in it, or the sound of his name. The sound of a bell may call up the idea of [p. 174] dinner. The outside world also is to them in large part a multitude of definite percepts. They feel the environment as trees, sticks, stones, chairs, tables, letters, words, etc. I have called such definite presentations 'free ideas' to distinguish them from the vague presentations such as atmospheric pressure, the feeling of malaise, of the position of one's body when falling, etc. It is such 'free ideas' which compose the substance of thought and which lead us to perhaps the majority of the different acts we perform, though we do, of course, react to the vaguer sort as well. I saw definitely in writing the last sentence the words 'majority of the different acts' and thought 'we perform' and so wrote it. I see a bill and so take check book and pen and write. I think of the cold outside and so put on an overcoat. This mental function 'having free ideas,' gives the possibility of learning to meet situations properly by thinking about them, by being reminded of some property of the fact before us or some element therein.
We can divide all learning into (1) learning by trial and accidental success, by the strengthening of the connections between the sense-impressions representing the situation and the acts - or impulses and acts - representing our successful response to it and by the inhibition of similar
connections with unsuccessful responses; (2) learning by imitation, where the mere performance by another of a certain act in a certain situation leads us to do the same; and (3) learning by ideas, where the situation calls up some idea (or ideas) which then arouses the act or in some way modifies it.
The last method of learning has obviously been the means of practically all the advances in civilization. The evidence quoted a paragraph or so back from the Experimental Study shows the typical mammalian mind to be one which [p. 175] rarely or never learns in this fashion. The present study of the primates has been a comparative study with two main questions in view: (1) How do the monkeys vary from the other mammals in the general mental functions revealed by their methods of learning? (2) How do they, on the other hand, vary from adult civilized human beings?
The experiments to be described seem, however, to be of value apart from the possibility of settling crucial questions by means of the evidence they give. To obtain exact accounts of what animals can learn by their own unaided efforts, by the example of their fellows or by the tuition of a trainer, and of how and how fast they learn in each case, seems highly desirable. I shall present the results in the manner which fits their consideration as arguments for or against some general hypotheses, but the naturalist or psychologist lacking the genetic interest may find an interest in them at their face value. I shall confine myself mainly to questions concerning the method of learning of the primates, and will discuss their sense-powers and unlearned reactions or instincts only in so far as is necessary to its comprehension.
It has been impossible for the author to make helpful use of the anecdotes and observations of naturalists and miscellaneous writers concerning monkey intelligence. The objections to such data pointed out in Chapter II, pp. 22-26, hold here. Moreover it is not practicable to sift out the true from the false or to interpret these random instances of animal behavior even if assuredly true. In the study of animal life the part is only clear in the light of the whole, and it is wiser to limit conclusions to such as are drawn from the constant and systematic study of a number of animals during a fairly long time. After a large enough body of such evidence has been accumulated we may be able to interpret random observations. [p. 176]
The subjects of the experiments were three South American monkeys of the genus Cebus. At the time of beginning the experiment No. 1 was about half grown, No. 2 was about one fourth full size and No. 3 was about half grown. No. 1 was under observation from November, 1899, to February, 1900; No. 2 and No. 3 from October, 1900, to February, 1901. No. 1 was during the period of experimentation decidedly tame, showing no fear whatever of my presence and little fear at being handled. He would handle and climb over me with no hesitation. No. 2 was timid, did not allow handling, but showed no fear of my presence and no phenomena that would differentiate his behavior in the experiments discussed from that of No. 1, save much greater caution in all respects. No. 3 also showed no fear at my presence. Any special individual traits that are of importance in connection with any of the observations will be mentioned in their proper places. No. 1 was kept until June, 1900, in my study in a cage 3 by 6 by 6 feet, and was left in the country till October, 1900. From October, 1900, all three were kept in a room 8 by 9 feet, in cages 6 feet tall by 3 long by 2.6 wide for Nos. 1 and 2, 3 feet by 3 feet by 20 inches for No. 3. I studied their behavior in learning to get into boxes, the doors to which could be opened by operating some mechanical contrivance, in learning to obtain food by other simple acts, in learning to discriminate between two signals, that is, to respond to each by a different act, and in their general life.
Following the order of the 'Animal Intelligence,' I shall first recount the observations of the way the monkeys learned, solely by their own unaided efforts, to operate simple mechanical contrivances.
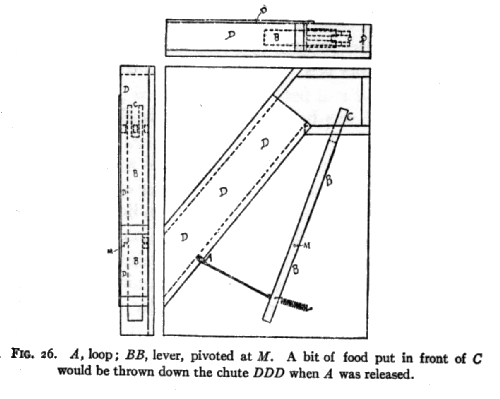
Besides a number of boxes such as were used with the dogs and cats (see illustration on P. 30), I tried a variety [p. 177] of arrangements which could be set up beside a cage, and which would, when some simple mechanism was set in action, throw a bit of food into the cage. Figure 26 shows one of these. See description of QQ (ff) on page 182.
APPARATUS
The different mechanisms which I used were the following: --
Box BB (O at back) was about 2o by 14 by 12 inches with a door in the front which was held by a bolt to which was tied a string. This string ran up the front of the box outside, over a pulley, across the top, and over another pulley down into the box, where it ended in a loop of wire.
Box MM (bolt) was the same as BB but with no string and loop attachment to the bolt.
Box CC (single bar) was a box of the same size as BB. [p. 178]
The door was held by a bar about 3 by 1 by 5 inches which swung on a nail at the left side.
Box CCC (double bar) was CC with a second similar bar on the right side of the door.
Box NN (hook) was a box about the size of BB with its door held by an ordinary hook on the left side which hooked through an eyelet screwed into the door.
Box NNN was NN with the hook on the right instead of the left side.
Box NNNN was box NN with two hooks, one on each side.
Apparatus 00 (string box) consisted of a square box tied to a string, which formed a loop running over a pulley by the cage and a pulley outside, so that pulling on the under string would bring the box to the cage. In each experiment the box was first pulled back to a distance Of 2 feet 3 inches from the cage, and a piece of banana put in it. The monkey could, of course, secure the banana by pulling the box near enough.
Apparatus 000 was the same as 00, with the box tied to the upper string, so that the upper string had to be pulled instead of the lower.
Box PP was about the size of BB. Its door was held by a large string securely fastened at the right, passing across the front of the door and ending in a loop which was put over a nail on the box at the left of the door. By pulling the string off the nail the door could be opened.
Box RR (wood plug) was a box about the size of BB. The door was held by a string at its top, which passed up over the front and top to the rear, where it was fastened to a wooden plug which was inserted in a hole in the top of the box. When the plug was pulled out of the hole, the door would fall open. [p. 179]
Box SS (triple; wood-plug, hook and bar) was a box about the size of BB. To open the door, a bar had to be pushed around, a hook unhooked and a plug removed from a hole in the top of the box.
Box TT (nail plug) was 14 by 10 by 10 inches with a door 5.5 by 10 on the right side of the front, the rest of the front being barred up. The door was hinged at the bottom and fastened at its top to a wire which was fastened to a nail 2.5 inches long, which, when inserted in a hole 0.25 inches in diameter at the back of the top of the box, held the door dosed. By drawing out this nail and pulling the door the animal could open the door.
Box VV (plug at side) was a box about 18 by 10 by 10, the door held by a plug passing through a hole in the side of the box. When the plug was pulled out, the door could be pushed inward.
Box W (loop) was 17 by 10 by 10 inches with a door 5 by 9 at the left side of its front hinged at the bottom. The door was prevented from falling inward by a wire stretched behind it. It was prevented from falling outward by a wire firmly fastened at the right side and held by a loop over a nail at the left. By pulling the loop outward and to the left it could be freed from the nail. The door could then be pulled open.
Box WW (bar inside) was 16 by 14 by 10 inches with a door 4 by 11 at the left of its front hinged at the bottom. The door could be pushed in or pulled out when a bar on its inside was lifted out of a latch. The bar was accessible from the outside through an opening in the front of the box. It had to be lifted to a height of 1.5 inches (an angle of about 30°).
Box XX (bar outside) was about 13 by 11 by 10 inches with a door 7 by 8 on the left side of the front. The door [p. 180] was held in place by a bar swinging on a nail at the top, with its other end resting in a latch at the left side of the box. By pushing this up through an angle Of 45° the door could be opened.
Box YY (push bar) was a box 16 by 8 by 12 inches with a door at the left of its front. The door was held by a brass bar which swung down in front of an L-shaped piece of steel fastened to the inside of the door. This brass bar was hung on a pivot at its center and the other end attached to a bar of wood the other end of this bar projected through a hole at the right side of the box. By pushing this bar in about an inch the door could be opened.
Box LL (triple nail plug, hook and bar) was a box 10 by 10 by 13 with a door 3 by 8.5 at the left side. The door could be opened only after (1) a nail plug had been removed from a hole in the back of the top of the box as in TT, (2) a hook in the door had been unhooked, and (3) a bar on the left side had been turned from a horizontal to a vertical position.
Box Alpha (catch at back) was 11 by 10 by 15 with the door (4 by 4) in the left side of its front. The door was held by a bolt, which, when let down, held in a catch on the inside of the door. A string fastened to the bolt ran across to the back of the box and through a hole to the outside. There it ended in a piece of wood 2.5 by 1 by .25 inches. When this piece of wood was pulled, the bolt went up and the door fell open.
Box Beta was the same as NN except in size. It was 10 by 10 by 13 inches.
Box KK (triple; bolt, side plug, and knob) was a box 16 by 9 by 11 with a door at the left side of the front. The door was held by a bolt on the right side, a wooden plug stuck through a hole in the box on its left side and a nail [p. 181] which held in a catch at its top. This nail was fastened to a wooden knob (1 by 5 by .375) which lay in a depression at the top of the box. Only when the bolt had been drawn and the plug and knob pulled, could the door be opened.
Box Gamma (wind) was 10 by 10 by 13 inches with its door held by a wire fastened at the top and wound three times about a screw eye in the top of the box. By unwinding the wire the door could be opened.
Box Delta (Push back) was 12 by 11 by 10 inches. Its door was held by a wooden bar projecting from the right two inches in front of it. This bar was so arranged that it could be pushed or pulled toward the right, allowing the door to fall open. It could not be swung up or down.
Box Epsilon (lever or push down) was 12 by 9 by 5 inches'. At the right side of its front was a hole 1 inch broad by ii inches up and down. Across this hole on the inside of the box was a strip of brass, the end of one bar of a lever. If this strip was depressed 1/8 of an inch, the door at the extreme left would be opened by a spring.
Box Zeta (side plug) was 12 by 11 by 10 inches. Its door was held by a round bar of wood put through a hoop of steel at the left side of the box. This bar was loose and could easily be pulled out, allowing the door to be opened.
Box Theta was the same as KK except that the door could be opened as soon as the bolt alone was pulled or pushed up.
Box Eta was like Alpha save that the object at the back of the box to be pulled was a brass ring.
Apparatus QQ (chute) consisted of a lever mechanism so arranged that by pushing in a bar of wood 1/4 to 1/2 an inch, a piece of banana would be thrown down a chute into the cage. The apparatus was placed outside the cage in such a [p. 182] way that it could be easily reached by the monkey's arm through the wire netting.
QQ (a) was of the same general plan. By turning a handle through 270° food could be obtained.
QQ (b) was like QQ (a) except that 21 full revolutions of the handle in one direction were necessary to cause the food to drop down.
QQ (c) was a chute apparatus so arranged as to work when a nail was pulled out of a hole.
QQ (d) was arranged to work at a sharp pull upon a brass ring hanging to it.
QQ (e) was arranged to work when a hook was unhooked.
QQ (f) was arranged to work when a loop at the end of a string was pulled off from a nail.
QQ (ff) was QQ (f) with a stiff wire loop instead of a loop of string.
EXPERIMENTS ON THE ABILITIES OF THE MONKEYS TO LEARN WITHOUT TUITION
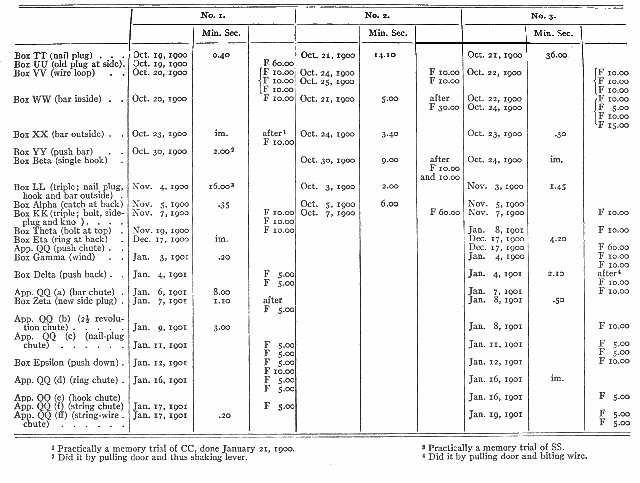
I will describe a few of the experiments with No. 1 as samples and then present the rest in the form of a table. No. 1 was tried first in BB (O at back) on January 17, 1900 being put inside. He opened the box by pulling up the string just above the bolt. His times were .05, 1.38, 6.00, 1.00, 10, .05, .05. He was not easily handled at this time, so I changed the experiment to the form adopted in future experiments. I put the food inside and left the animal to open the door from the outside. He pulled the string up within 10 seconds each time out of 10 trials.
I then tried him in MM (bolt). He failed in 15. 1 then (January 18th) tried him in CC (single bar outside.) He got in in 36.00 minutes; he did not succeed a second time [p. 183] that night, but in the morning the box was open. His times thenceforth were 20, 10, 16, 25 and on January 19th, 40, 5, 12, 8, 5, 5, 5 seconds.
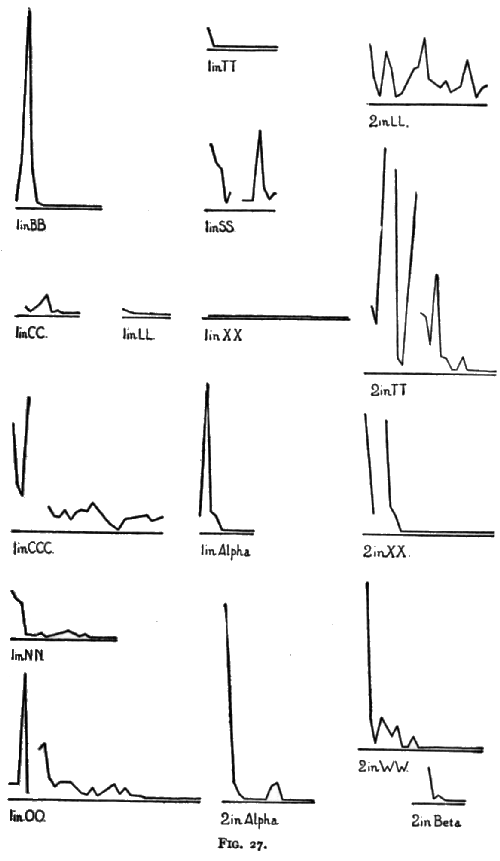
I then tried him (January 21, 1900) in CCC (double bar). He did it at first by pushing the old bar and then pulling at the door until he worked the second bar gradually around. Later he at times pushed the second bar. The times taken are shown in the time-curve. I then (January 25th) tried him in NN (hook). See time-curves on page 185. I then (January 27th) tried him in NNN (hook on other side). He opened it in 6, 12 and 4 seconds in the first three trials. I then (20 minutes later) tried him with NNNN (double hook). He opened the door in 12, 10, 6 and 6 seconds. I then (January 27th) tried him with PP (string across). He failed in 10. I then (February 21st) tried him with apparatus 00 (string box). For his progress as shown by the times taken see the time-curve. His progress is also shown in the decrease of the useless pullings at the wrong string. There were none in the 9th trial, 14th, 15th, 16th, 18th, 24th, and following trials.
No. 1 was then (February 24th) tried with 000 (string box with box on upper string). No. i succeeded in 2.20, then failed in 10.00 The rest of the experiment will be described under imitation.
He was next tried (March 24th) with apparatus QQ (chute). He failed in 10.00, though he played with the apparatus much of the time. Other experiments were with box RR (wood-plug) (April 5th). He failed in 10.00. After he had, in a manner to be described later, come to succeed with RR, he was tried in box SS (triple; wood-plug, hook and bar) (April 18th); see time-curve. No more experiments of this nature were tried until October, 1900.
The rest of the experiments with No. 1 and all those with [p. 184]
No. 2 and No. 3 may best be enumerated in the form of a table. (See Table 9 on page 187.) It will show briefly the range of performances which the unaided efforts of the animals can cope with. It will also give the order in which each animal experienced them. F means that the animal failed to succeed. The figures are minutes and seconds, and represent the time taken in the first trial or the total time taken without success where there is an F. In cases where the animal failed in say 10 minutes, but in a later trial succeeded, say in 2.40, the record will be 2.40 after 10 F. There are separate columns for all three animals, headed No. 1 No. 2 and No. 3. Im. stands for a practically immediate success.
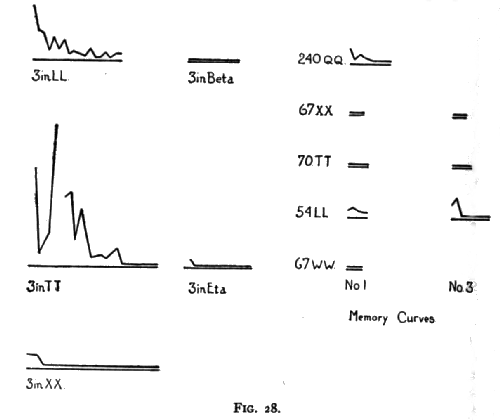
The curves on pages 185 and 186 (Figs. 27 and 28) show the progress of the formation of the associations in those cases where the animal was given repeated trials, with, however, nothing to guide him but his own unaided efforts. Each millimeter on the abscissa represents one trial and each millimeter on the ordinate represents 10 seconds, the ordinates representing the time taken by the animal to open the box. A break in the curve, or an absence of the curve at the beginning of the base-line represents cases where the animal failed in 10 minutes or took a very long time to get out.
In discussing these facts we may first of all clear our way of one popular explanation, that this learning was due to 'reasoning.' If we use the word reasoning in its technical psychological meaning as the function of reaching conclusions by the perception of relations, comparison and inference, if we think of the mental content involved as feelings of relation, perceptions of similarity, general and abstract notions and judgments, we find no evidence of reasoning [p. 185]
[p. 186] in the behavior of the monkeys toward the mechanisms used. And this fact nullifies the arguments for reasoning in their case as it did in the case of the dogs and cats. The argument that successful dealings with mechanical contrivances imply that the animals reasoned out the properties
of the mechanisms, is destroyed when we find mere selection from their general instinctive activities sufficient to cause success with bars, hooks, loops, etc. There is also in the case of the monkeys, as in that of the other mammals, positive evidence of the absence of any general function of reasoning. We shall find that at least very many simple acts were not learned by the monkeys in spite of their having seen me perform them again and again; that the same holds true of many simple acts which they saw other monkeys do, or were put through by me. We shall find that after having [p. 187]
[p. 188] abundant opportunity to realize that one signal meant food at the bottom of the cage and another none, a monkey would not act from the obvious inference and consistently stay up or go down as the case might be, but would make errors such as would be natural if he acted under the growing influence of an association between sense-impression and impulse or sense-impression and idea, but quite incomprehensible if he had compared the two signals and made a definite inference. We shall find that, after experience with several pairs of signals, the monkeys yet failed, when a new pair was used, to do the obvious thing to a rational mind; viz., to compare the two, think which meant food, and act on the knowledge directly.
The methods one has to take to get them to do anything, their general conduct in becoming tame and in the experiments throughout, confirm these conclusions. The following particular phenomena are samples of the many which are inconsistent with the presence of reasoning as a general function. No. 1 had learned to open a door by pushing a bar around from a horizontal to a vertical position. The same box was then fitted with two bars. He turned the first bar round thirteen times before attempting to push the other bar around. In box LL all three monkeys would in the early trials do one or two of the acts over and over after they had once done them. No. 1, who had learned to pull a loop of wire off from a nail, failed thereafter to pull off a similar loop made of string. No. 1 and No. 3 had learned to poke their left hands through the cage for me to take and operate a chute with. It was extremely difficult to get either of them to put his right hand through or even to let me take it and pull it through.
A negative answer to the question "Do the monkeys reason?" thus seems inevitable, but I do not attach to [p. 189] the question an importance commensurate with the part it has played historically in animal psychology. For I think it can be shown, and I hope in a later monograph to show, that reasoning is probably but one secondary result of the general function of having free ideas in great numbers, one product of a type of brain which works in great detail, not in gross associations. The denial of reasoning need not mean, and does not to my mind, any denial of continuity between animal and human mentality or any denial that the monkeys are mentally nearer relatives to man than are the other mammals.
So much for supererogatory explanation. Let us now turn to a more definite and fruitful treatment of these records.
The difference between these records and those of the chicks, cats and dogs given on pages 39-65 passim is undeniable. Whereas the latter were practically unanimous, save in the cases of the very easiest performances, in showing a process of gradual learning by a gradual elimination of unsuccessful movements, and a gradual reënforcement of the successful one, these are unanimous, save in the very hardest, in showing a process of sudden acquisition by a rapid, often apparently instantaneous, abandonment of the unsuccessful movements and a selection of the appropriate one which rivals in suddenness the selections made by human beings in similar performances. It is natural to infer that the monkeys who suddenly replace much general pulling and clawing by a single definite pull at a hook or bar have an idea of the hook or bar and of the movement they make. The rate of their progress is so different from that of the cats and dogs that we cannot help imagining as the cause of it a totally different mental function, namely, free ideas instead of vague sense-impres- [p. 190] sions and impulses. But our interpretation of these result should not be too hasty. We must first consider sever other Possible explanations of the rapidity of learning by the monkeys before jumping to the conclusion that the forces which bring about the sudden formation of associations in human beings are present.
First of all it might be that the difference was due to the superiority of the monkeys in clear detailed vision. It might be that in given situations where associations were to be formed on the basis of smells, the cats and dogs would show similar rapid learning. There might be, that is, no general difference in type of mental functioning, but only a special difference in the field in which the function worked. This question can be answered by an investigation of the process of forming associations in connection with smells by dogs and cats. Such an investigation will, I hope, soon be carried on in the Columbia Laboratory by Mr. Davis.[4]
Secondly, it might be that the superior mobility and more detailed and definite movements of the monkeys' hands might have caused the difference. The slowness in the case of the dogs and cats might be at least in part the result of difficulty in executing movements, not in intending them. This difficulty in execution is a matter that cannot be readily estimated, but the movements made by the cats and dogs would not on their face value seem to be hard. They were mostly common to the animals' ordinary life. At the same time there were certain movements (e.g. depressing the lever) which were much more quickly associated with their respective situations by the cats than others were, and if we could suppose that all the movements learned by the monkeys were comparable to these few, it would detract [p. 191] from the necessity of seeking some general mental difference as the explanation of the difference in the results.
In the third place it may be said by some that no comparison of the monkeys with dogs and cats is valid, since the former animals got out of boxes while the latter got in. It may be supposed that the instinctive response to confinement includes an agitation which precludes anything save vague unregulated behavior. Professor Wesley Mills has made such a suggestion in referring to the 'Animal Intelligence' in the Psychological Review, May, 1899. In the July number of the same journal I tried to show that there was no solid evidence of such a harmful agitation. Nor can we be at all sure that agitation when present does not rather quicken the wits of animals. It often seems to. However I should, of course, allow that for purposes of comparison it would be better to have the circumstances identical. And I should welcome any antagonist who should, by making experiments with kittens after the fashion of these with the monkeys, show that they did learn as suddenly as the latter.
Again we know that, whereas the times taken by a cat in a box to get out are inversely proportional to the strength of the association, inasmuch as they represent fairly the amount of its efforts, on the other hand, the times taken by a monkey to get in represent the amounts of his efforts plus the amount of time in which he is not trying to get in. It may be said therefore that the time records of the monkeys prove nothing, - that a record of four minutes may mean thirty seconds of effort and three minutes thirty seconds of sleep, that one minute may really represent twice as much effort. As a matter of fact this objection would occasionally hold against some single record. The earliest times and the occasional long times amongst very short ones are likely [p. 192] to be too long. The first fact makes the curves have too great a drop at the start, making them seem cases of too sudden learning, but the second fact makes the learning seem indefinite when it really is not. And in the long run the times taken do represent fairly well the amount of effort. I carefully recorded the amount of actual effort in a number of cases and the story it tells concerning the mental processes involved is the same as that told by the time-curves.
Still another explanation is this: The monkeys learn quickly, it is true, but not quickly enough for us to suppose the presence of ideas, or the formation of associations among them. For if there were such ideas, they should in the complex acts do even better than they did. The explanation then is a high degree of facility in the formation of associations of just the same kind as we found in the chicks, dogs and cats.
Such an explanation we could hardly disapprove in any case. No one can from objective evidence set up a standard of speed of learning below which all shall be learning without ideas and above which all shall be learning by ideas. We should not expect any hard and fast demarcation.
This whole matter of the rate of learning should be studied in the light of other facts of behavior. My own judgment, if I had nothing but these time-curves to rely on, would be that there was in them an appearance of learning by ideas which, while possibly explicable by the finer vision and freer movements of the monkey in connection with ordinary mammalian mentality, made it worth while to look farther into their behavior. This we may now do.
What leads the lay mind to attribute superior mental gifts to an animal is not so much the rate of learning as the amount learned. The monkeys obviously form more [p. 193] associations and associations in a greater variety than do the other mammals. The improved rate assists, but another cause of this greater number of associations is the general physical activity of the monkeys, their constant movements of the hands, their instinctive curiosity or tendency to fool with all sorts of objects, to enjoy having sense-impressions, to form associations because of the resulting sound or sight. These mental characteristics are of a high degree of importance from the comparative point of view, but they cannot be used to prove that the monkeys have free ideas, for a large number of associations may be acquired after the purely animal fashion.
What is of more importance is the actual behavior of the animals in connection with the boxes. First of all, as has been stated, all the monkey's movements are more definite, he seems not merely to pull, but to pull at, not merely to poke, but to push at. He seems, even in his general random play, to go here and there, pick up this, examine the other, etc., more from having the idea strike him than from feeling like doing it. He seems more like a man at the breakfast table than like a man in a fight. Still this appearance may be quite specious, and I think it is likely to lead us to read ideational life into his behavior if we are not cautious. It may be simply general activity of the same sort as the narrower activities of the cat or dog.
In the second place the monkeys often make special movements with a directness which reminds one unavoidably of human actions guided by ideas. For instance, No. 1 escaped from his cage one day and went directly across the room to a table where lay a half of a banana which was in a very inconspicuous place. It seemed as if he had observed the banana and acted with the idea of its position fully in mind. Again, on failing to pull a hook out, No. 1 im- [p. 194] mediately applied his teeth, though he had before always pulled it out with his hand. So again with a plug. It may be that there is a special inborn tendency to bite at objects pulled unsuccessfully. If not, the act would seem to show the presence of the idea 'get thing out' or 'thing come out' and associated with it the impulse to use the teeth. We shall see later, however, that in certain other circumstances where we should expect ideas to be present and result in acts they do not.
The fact is that those features in the behavior of the monkeys in forming associations between the sight of a box and the act needed to open it which remind us of learning by ideas may also be possibly explained by general activity and curiosity, the free use of the hand, and superior quickness in forming associations of the animal sort. We must have recourse to more crucial tests or at least seek evidence from a number of different kinds of mental performances. The first of these will naturally be their behavior toward these same mechanisms after a long time-interval.
THE PERMANENCE OF ASSOCIATIONS IN TIRE CASE OF MECHANISMS
My records are too few and in all but one case after too short an interval to be decisive on the point of abrupt transition from failure to success such as would characterize an animal in whose mind arose the idea of a certain part of the mechanism as the thing to be attacked or of a certain movement as the fit one. The animals are all under observation in the Columbia Laboratory, however, and I trust that later satisfactory tests may be made. No. 2 was not included in the tests because he was either unwell or had become very shy of the boxes, entering them even [p. 195] when the door was left open only after great delay. The time-curves for the experiments performed will be found on page 186 among the others. The figures beside each pair represent the number of days without practice.
The records show a decided superiority to those of the cats and dogs. Although the number of trials in the original tests were in general fewer in the case of the monkeys, the retention of the association is complete in 6 cases out of 8 and is practically so in one case where the interval was 8 months.
EXPERIMENTS ON THE DISCRIMINATION OF SIGNALS
My experiments on discrimination were of the following general type: I got the animal into the habit of reacting to a certain signal (a sound, movement, posture, visual presentation or what not) by some well-defined act. In the cases to be described this act was to come down from his customary positions about the top of the cage, to a place at the bottom. I then would give him a bit of food. When this habit was wholly or partly formed, I would begin to mix with that signal another signal enough like it so that the animal would respond in the same manner. In the cases where I gave this signal I would not feed him. I could then determine whether the animal did discriminate or not, and his progress toward perfect discrimination in case he did. If an animal responds indiscriminately to both signals (that is, does not learn to disregard the 'no food' signal) it is well to test him by using two somewhat similar signals, after one of which you feed him at one place and after the other of which you feed him at a different place.
If the animal profits by his training by acquiring ideas of [p. 196] the two signals and associates with them ideas of 'food I and 'no food,' 'go down' and 'stay still,' and uses these ideas to control his conduct, he will, we have a right to expect, change suddenly from total failure to differentiate the signals to total success. He will or won't have the ideas, and will behave accordingly. The same result could, of course, be brought about by very rapid association of the new signal with the act of keeping still, a very rapid inhibition of the act of going down in response to it by virtue of the lack of any pleasure from doing so.
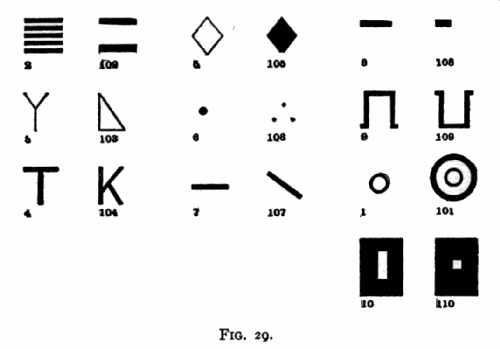
For convenience I shall call the signals after which food was given yes signals and those after which food was not given no signals. Signals not described in the text are shown in Fig. 29, below. The progress of the monkeys in
discriminating is shown by Figs. 30 and 31, on pages 199 and 201. In Figs. 30 and 31 every millimeter along the horizontal or base line represents 10 trials with the signal. The heights of the black surface represent the percentages of wrong responses, 10 mm. meaning 100 per cent of [p. 197] incorrect responses. Thus the first figure of the set, Left hand, a, presents the following record: First 10 trials, all wrong; of next 10, 7 wrong; of next 10, 6 wrong; of next 10, 7; of the next, 9; of the next, 9; of the next, 4; Of the next, none; of the next, 3; of the next, 2, and then 70 trials without an error.
I will describe some of the experiments in detail and then discuss the graphic presentation of them all.
EXPERIMENTS WITH No. 1
Having developed in No. 1 the habit of coming down to the bottom of his cage to get a bit of food when he saw me reach out and take such a bit from my desk, 1 tested his ability to discriminate by beginning to use now one hand, now the other, feeding him only when I used the left. 1 also used different sets of words, namely, 'I will give some food' and 'They shall not have any.' It will be seen later that he probably reacted only to the difference of the hands. The experiment is similar to that described on pages 129 and 130 of Chapter II. At the beginning, it should be remembered, No. 1 would come down whichever hand was used, no matter what was said, except in the occasional cases where he was so occupied with some other pursuit as to be evidently inattentive. He did come to associate the act of going down with the one signal and the act of staying still or continuing his ordinary movements with the other signal. His progress in learning to do so is best seen in the curves of his errors. To the 'yes' signal he responded correctly, except for the occasional lapses which I just mentioned, from the start and throughout. With the 'no' signal his errors were as shown in Fig. 30, a. The break in the curve at 110 and 120 is probably not significant [p. 198] of an actual retrograde as the trials concerned followed an eight days' cessation of the experiments.
I next tried No. 1 with an apparatus exposing sometimes a card with a diamond-shaped piece of buff-colored paper, on it and sometimes a card with a similar black piece. The black piece was three fourths of an inch farther behind the opening than the other. The light color was the 'yes' signal. The error curves for both signals are given, as No. 1 at the beginning of the experiment did not go down always (Fig. 30, b and b1 ).
I next tried No. 1 with the same apparatus but exposing cards with YES and N in place of the buff and black diamonds. The record of the errors is given in Fig. 30, c and c1. At the start he came down halfway very often. This I arbitrarily scored as an error no matter which signal it was in response to. It should not be supposed that these curves represent two totally new associations. It seems likely that the monkey reacted to the position of the N card in the apparatus (the same as that of the black diamond card) rather than to the shape of the letters. On putting the black diamond in front he was much confused.
I next gave No. 1 the chance to form the habits of coming down when I rapped my pencil against the table twice and of staying where he was when I rapped with it once. He had go trials of each signal but failed to give evidence of any different associations in the two cases.
Experiments of this sort were discontinued in the summer. In October I tried No. 1 with the right and left hand experiment, he being in a new room and cage, and I being seated in a different situation. He came down at both signals and failed to make any ascertainable progress with the no signal in 80 trials. (October 20-24.)
I then tried him with the black and buff diamonds, the [p. 199]
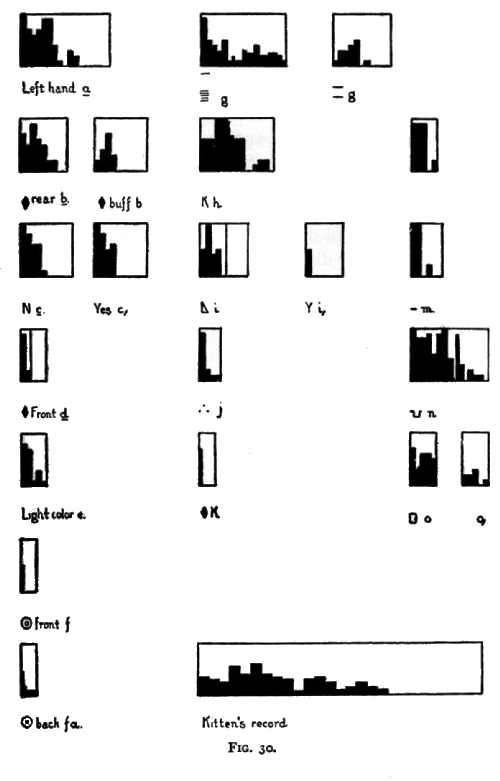
[p. 200] black being in front (October 25-29). The reaction to the 'yes' signal was perfect from the start. The progress with the 'no' signal is shown in Fig. 30, d.
I then tried him with an apparatus externally of different size, shape and color from that so far used, showing as the 'yes' signal a brown card and as the 'no' signal a white and gold card one half inch farther back in the apparatus. The 'yes' signal was practically perfect from the start. His progress with the 'no' signal is shown in Fig. 3o, e.
I then tried a still different arrangement for exposure, to which, however, he did not give uniform attention.
I then tried cards 1 and 101, 101 being in front and 1 in back. 1 was the 'yes' signal. 'Yes' responses were perfect from the start. For 'no' responses see Fig. 30, f. I then put the 'Yes' signal in front and the 'no' signal behind. 'Yes' responses perfect; for 'no' responses see Fig. 30, f, a.
From now on I arranged the exposures in such a way that there was no difference between the 'yes' and 'no' signals in distance or surroundings.
The following list shows the dates, signals used, and the figures on page 199 presenting the results. Where there is only one figure drawn, it refers to progress with the 'no' signal, the 'yes' signal being practically perfect from the start.
[p. 201]
Fig. 29 gives facsimiles of the different signals reduced to one sixth their actual size. The drawing of 101 is not accurate, the outer ring being too thick.
EXPERIMENTS WITH No. 2
I first secured the partial formation of the habit of coming down when I took a bit of food in my hand. I then used the apparatus for exposing cards, YES in front being the 'yes' signal and a circle at the back being the 'no' signal. I gave No. 2 25 trials with the 'yes' signal and then began a regular
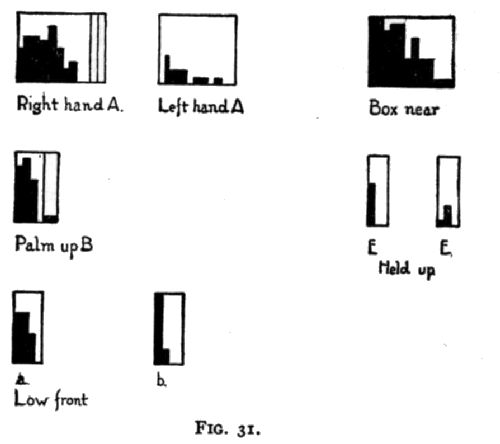
experiment similar to those described. After about go trials (November 9-12, 1900 ) there was no progress toward differentiation of response, and it was evident from No. 2's behavior that he was reacting solely to the movements of my hand. So I abandoned the exposing apparatus and used (November 11-13, 1900) as the 'yes' signal the act of taking the food with my left hand from a pile on the front of the box and for the' no' signal the act of taking food with my right hand from a pile 4 inches behind that just mentioned. [p. 202] No. 2 did come to differentiate these two signals. The record of his progress is given in Fig. 31 by A and A 1.
I then made a second attempt with the exposing apparatus, using cards 2 and 102 (November 6, 14-21). No. 2 did react to my movements in pulling the string but in over 100 trials made no progress in the direction of a differential reaction to the 'no' signal. I then tried feeding him at each signal, feeding him at the bottom of the cage as usual when I gave the 'yes' signal and at the top when I gave the 'no' signal. After a hundred trials with the 'no' signal there was no progress.
I then abandoned again the exposing apparatus and used as signals the ordinary act of taking food with my left hand (yes) and the act of moving my left arm from my right side round diagonally (swinging it on my elbow as a center) and holding the hand, after taking the food, palm up (no) (November 26, 27, 1900). No. 2 did come to differentiate these signals. His progress is given in the diagram in Fig. 31 entitled 'Palm up' (B).
I next used (November 27, 1900) as the 'yes' signal the same act as before and for the 'no' signal the act of holding the food just in front of the box about four inches below the edge. No. 2's progress is shown in Fig. 31 in the diagram entitled 'low front' (C and C 1 ).
I next used (November 27-30) the same movement for both 'yes' and 'no' signals save that as the 'yes' signal I took the food from a brown pasteboard box 3 by 3 by 0.5, and as the 'no' signal I took it from a white crockery cover two inches in diameter and three eighths of an inch high which was beside the box but three inches nearer me. No. 2'S progress is shown in Fig- 31 in the diagram entitled 'Box near ' (D).
I next used for the 'yes' signal the familiar act and for the [p. 203] 'no' signal the act of holding the food six inches above the box instead of a quarter or a half an inch. The progress is shown in Fig. 31, E and E 1. I then tried taking the food from a saucer off the front of the box for the 'yes' signal and from a small box at the back for the 'no' signal. 'Yes' was perfect from the start (10 trials given). 'No' was right once, then wrong once, then right for the remaining eight.
EXPERIMENTS WITH No. 3
No. 3 was kept in a cage not half so big as those of 1 and 2. Perhaps because of the hindrance this fact offered to forming the habit of reacting in some definite way to 'yes' signals, perhaps because of the fact that I did not try hand movements as signals, there was no successful discrimination by No. 3 of the yellow from the black diamond or of a card with YES from a card with a circle on it. I tried climbing up to a particular spot as the response to the 'yes' signal and stay in still as the response to the 'no' signal. I also tried instead of the latter a different act, in which case the animal was fed after both signals but in different places. In the latter case No. 3 made some progress, but for practical reasons I postponed experiments with him. Circumstances have made it necessary to postpone such experiments indefinitely.
PERMANENCE OF THE ABILITY TO DISCRIMINATE
No. 1 and No. 2 were tried again after intervals of 33 to 48 days. The results of these trials are shown in Fig-32. Here every millimeter along the base line represents one trial with the 'no' signal (the 'yes' signals were practically perfect), and failure is represented by a column 10 m.m. high while [p. 204] success is represented by the absence of any column. Thus the first record reads, "No. 1 with signal 104 after 40 days

made 5 failures, then 2 successes, then 1 failure, then 1 success, then 3 failures, then 1 success, then 1 failure, 1 then 3 successes, then 1 failure, then 10 successes." The third record (106; 40 days) reads, "perfect success in ten trials."
DISCUSSION OF RESULTS
The results of all these discrimination experiments emphasize the rapidity of formation of associations amongst the monkeys, which appeared in their behavior toward the mechanisms. The suddenness of the change in many cases is immediately suggestive of human performances. If all the records were like c, f, h, i, j, k, 1, m, B, E, and memory trials 103, A, B, and C, one would have to credit the animals with either marvelous rapidity in forming associations of the purely animal sort or concede that from all the objective evidence at hand they were shown to learn as human beings would. One would have to suppose that they had clear [p. 205] ideas of the signals and clean-cut associations with those ideas. The other records check such a conclusion.
In studying the figures we should remember that occasional mistakes, say 1 in 10 trials, are probably not significant of incomplete learning but of inattention or of precipitate action before the shutter had fairly exposed the card. We must not expect that a monkey who totally fails to discriminate will always respond wrongly to the 'no' signal, or that a monkey who has come to discriminate perfectly will always respond rightly. A sudden drop from an average high level of error to an average low level will signify sudden learning. Where the failure was on the first trial of a series a few hours or a day removed from the last series, I have generally represented the fact not by a column i mm. high and i mm. broad, but by a single 10 mm. perpendicular. See i and A. Such cases represent probably the failure of the animal to keep his learning permanent rather than any general inability to discriminate.
K was to some extent a memory trial of d (after over half a year).
The experiment with 10 and 110 is noteworthy. Although, as can be seen from the figures, the difference is obvious to one looking at the white part of the figure, it is not so to one looking at the black part. No. 1 failed to improve appreciably in fifty trials, probably because his previous experience had gotten him into the habit of attending to the black lines.
Before arguing from the suddenness of the change from failure to success we have to consider one possibility that I have not mentioned, and in fact for the sake of clearness in presentation have rather concealed. It is that the sudden change in the records, which report only whether the animal did or did not go down, may represent a more gradual [p. 206] change in the animal's mind, a gradual weakening of the impulse to go down which makes him feel less and less inclined to go down, though still doing so, until this weakening reaches a sort of saturation point and stops the action. There were in their behavior some phenomena which might witness to such a process, but their interpretation is so dependent on the subjective attitude and prepossessions of the observer that I prefer not to draw any conclusions from them. On the other hand, records c, g, n, A and D seem to show that gradual changes can be paralleled by changes in the percentage of failures.
In the statement of conclusions I shall represent what would be the effect on our theory of the matter in both cases, (1) taking the records to be fairly perfect parallels of the process, and (2) taking them to be the records of the summation points of a process not shown with surety in any measurable objective facts. But I shall leave to future workers the task of determining which case is the true one.
If we judge by the objective records themselves, we may still choose between two views. (1) We may say that the monkeys did come to have ideas of the acts of going down to the bottom of the cage and of staying still, and that their learning represented the association of the sense-impressions of the two signals, one with each of these ideas, or possibly their association with two other ideas (of being fed, and of not being fed), and through them with the acts. Or (2) we may say that the monkeys had no such ideas, but merely by the common animal sort of association came to react in the profitable way to each signal.
If we take the first view, we must explain the failure of the animals to change suddenly in some of the experiments, must explain why, for instance, No. 1 in g should, after he had responded correctly to the 'no' signal for 27 trials out Of 30, [p. 207] fail in one trial out of four for a hundred or more trials. If the 2 7 successes were due to ideas, why was there regression ? If the animal came to respond by staying still on seeing the K (card 104), because that sight was associated with the idea of no food or the idea of staying still, why did he, in his memory trial, act sometimes rightly, sometimes wrongly, for eleven trials after his acting rightly twice. If he stayed still because the idea was aroused, why did he not stay still as soon as he had a few trials to remind him of the idea? It is easy, one may say, to see why, with a capacity to select movements and associate them with sense-presentations very quickly, in cases where habit provides only two movements for selection and where the sense-presentation is very dear and simple, an animal should practically at once be confirmed in the one act on an occasion when he does it with the sense-impression in the focus of attention. It is easy, therefore, to explain the sudden change in i, l, in, B, C and E. But our critic may add, "It is very hard to suppose that an animal that learned by connecting the sight of a card with the idea 'stay still' or the idea 'no food,' should be so long in making the connection as was the case in some of these experiments, should take 10, 20 or 40 trials to change from a high percentage of wrong to a high percentage of right reactions."
If we take the second view, we have to face the fact that many of the records are nothing like the single one we have for comparison, that of the kitten shown in Fig. 30, and that the appeal to a capacity to form animal associations very quickly seems like a far-fetched refuge from the other view rather than a natural interpretation. If we take the records to be summation points in a more gradual process, this difficulty is relieved.
If further investigation upheld the first view, we should [208] still not have a demonstration that the monkeys habitually did learn by getting percepts and images associated with sense-impressions, by having free ideas of the acts they performed; we should only have proved that they could under certain circumstances.
The circumstances in these experiments on discrimination were such as to form a most favorable case. The act of going down had been performed in all sorts of different connections and was likely to gain representation in ideational life; the experience 'bit of banana' had again been attended to as a part of very many different associations and so would be likely to develop into a definite idea.
These results then do not settle the choice between three theories: (1 a) that they were due to a general capacity for having ideas, (1 b) that they were due to ideas acquired by specially favoring circumstances, (2) that they were due to the common form of association, the association of an impulse to an act with a sense-impression rather roughly felt.
It would be of the utmost interest to duplicate these experiments with dogs, cats and other mammals and compare the records. Moreover, since we shall find (1 a) barred out by other experiments, it will be of great interest to test the monkeys with some other type of act than discrimination to see if, by giving the animal experience of the act and result involved in many different connections, we can get a rate of speed in the formation of a new association comparable to the rates in some of these cases.
Of course here, as in our previous section, the differences in the sense-powers of the monkeys from those of the kitten which I have tested with a similar experiment may have caused the difference in behavior. Focalized vision lends itself to delicate associations. Perhaps if one used the sense of smell, or if the dogs and cats could, preserving their same [p. 209] mental faculties in general, add the capacity for focalized vision, they would do as well as the monkeys.
EXPERIMENTS ON THE INFLUENCE OF TUITION
The general aim of these experiments was to ascertain whether the monkeys' actions were at all determined by the presence of free ideas and if so, to what extent. The question is, "Are the associations which experience leads them to form, associations between (1) the idea of an object and (2) the idea of an act or result and (3) the impulses and act itself, or are they merely associations between the sense-impression of the object and the impulse and act?" Can a monkey learn and does he commonly learn to do things, not by the mere selection of the act from amongst the acts done by him, but by getting some idea and then himself providing the act because it is associated in his mind with that idea. If a monkey feels an impulse to get into a box, sees his arm push a bar and sees a door fall open immediately thereafter and goes into the box enough times, he has every chance to form the association between the impulse to get into the box and the idea 'arm push bar,' provided he can have such an idea. If his general behavior is due to having ideas connected with and so causing his acts, he has had chance enough to form the association between the idea 'push at' and the act of pushing. If then a monkey forms an association leading to an act by being put through the act, we may expect that he has free ideas. And if he has free ideas in general in connection with his actions, we may expect him to so form associations. So also if a monkey shows a general capability to learn from seeing another monkey or a human being do a thing. A few isolated cases of imitation, however, might witness not to any general mental quality, [p. 210] but only to certain instincts or habits differing from others only in that the situation calling forth the act was the same act performed by another.
If the monkeys do not learn in these ways, we must, until other evidence appears, suppose them to be in general destitute of a life of free ideas, must regard their somewhat ambiguous behavior in learning by their own unaided efforts as of the same type as that of the dogs and cats, differing only in the respects mentioned on pages 190 and 191.
The general method of experimentation was to give monkeys who had failed of their own efforts to operate some simple mechanism, a chance to see me do it or see another monkey do it or to see and feel themselves do it, and then note any change in their behavior. The chief question is whether they succeed after such tuition when they have failed before it, but the presence of ideas would also be indicated if they attacked, though without success, the vital point in the mechanism when they had not done so before. On the other hand, mere success would not prove that the tuition had influenced them, for if they made a different movement or attacked a different spot, we could not attribute their behavior to getting ideas of the necessary act.
The results of the experiments as a whole are on their face value a trifle ambiguous, but they surely show that the monkeys in question had no considerable stock of ideas of the objects they dealt with or of the movements they made and were not in general capable of acquiring, from seeing me or one of their comrades attack a certain part of a mechanism and make a certain movements any ideas that were at all efficacious in guiding their conduct. They do not acquire or use ideas in anything that approaches the way human adults do. Whether the monkeys may not have some few ideas corresponding to habitual classes of objects and acts [p. 211] is a different question. Such may be present and function as the excitants of acts.
It is likely that this question could have been definitely solved if it had been possible for me to work with a larger number of animals. With enough subjects one could use the method mentioned on page 10 of Chapter II, of giving the animals tuition in acts which they would eventually do themselves without it, and then leaving them to their efforts, noting any differences in the way they learned from that in which other subjects who had no tuition learned the same acts. The chief of such differences to note would be differences in the time of their first trial, in the slope of the time-curve and in the number of useless acts.
It would also be possible to extend experiments of the type of the (on chair) experiment, where a subject is given first a certain time (calculated by the experimenter to be somewhat less than would be needed for the animal to hit upon the act) and if he does fail is then given certain tuition and then a second trial. The influence of the tuition is estimated by the presence or absence of cases where after tuition the act is done within the time.
There is nothing necessarily insoluble in the problem. Given ten or twenty monkeys that can be handled without any difficulty and it could be settled in a month.
With this general preface we may turn to the more special questions connected with the experiments on imitation of human acts and of the acts of other monkeys and on the formation of associations apart from the selection of impulses.
Imitation of Human Beings
It has been a common opinion that monkeys learned to do things from seeing them done by human beings. [p. 212]
We find anecdotes to that effect in fairly reputable authors.
Of course, such anecdotes might be true and still not prove that the animals learned to do things because they saw them done. The animal may have been taught in other ways to respond to the particular sights in question by the particular acts. Or it may have been in each case a coincidence.
If a monkey did actually form an association between a given situation and act by seeing some one respond to that situation by that act, it would be evidence of considerable importance concerning his general mental status, for it would go to show that he could and often did form associations between sense-impressions and ideas and between ideas and acts. Seeing some one turn a key in a lock might thus give him the idea of turning or moving the key, and this idea might arouse the act. However, the mere fact that a monkey does something which you have just done in his presence need not demonstrate or even render a bit more probable such a general mental condition. For he perhaps would have acted in just the same manner if you had offered him no model. If you put two toothpicks on a dish, take one and put it in your mouth, a monkey will do the same, not because he profits by your example, but because he instinctively puts nearly all small objects in his mouth. Because of their general activity, their instinctive impulses to grab, drop, bite, rub, carry, move about, turn over, etc., any novel object within their reach, their constant movement and assumption of all sorts of postures, the monkeys perform many acts like our own and simulate imitation to a far greater extent than other mammals.
Even if a monkey which has failed of itself to do a certain thing does it after you have shown him the act, there need be no reason to suppose that he is learning by imitation, [p. 213] forming an association between the sight of the object and. the act towards it through an idea gained from watching you. You may have caused his act simply by attracting his attention to the object. Perhaps if you had pointed at it or held it passively in your hand, yon would have brought to pass just the same action on his part. There are several cases among my records where an act which an animal failed totally to do of himself was done after I had so attracted his attention to the object concerned.
Throughout all the time that I had my monkeys under observation I never noticed in their general behavior any act which seemed due to genuine imitation of me or the other persons about. I also gave them special opportunities to show such by means of a number of experiments of the following type: where an animal failed by himself to get into some box or operate some mechanism, I would operate it in his presence a number of times and then give him a chance to profit by the tuition. His failure might be due to (1) the absence of instinctive impulses to make the movement in that situation, (2) to lack of precision in the movement, (3) to lack of force, or (4) to failure to notice and attack some special part of the mechanism. An instance of (1) was the failure to push away from them a bar which held a door; an instance of (a) was the failure to pull a wire loop off a nail; an instance of (2) or (3) was the failure to pull up a bolt; an instance of (4) was the failure to pull up an inside bar. Failures due to (3) occur rarely in the case of such mechanisms as were used in my investigations.
The general method of experiment was to make sure that the animal would not of itself perform a certain act in a certain situation, then to make sure that his failure could not be remedied by attracting his attention to the object, then to perform the act for him a number of times, letting him get [p. 214] each time the food which resulted, and finally to see whether, having failed before the tuition, he would succeed after it. This sounds very simple, but such experiments are hard to carry out satisfactorily. If you try the animal enough times by himself to make quite sure that he will not of himself hit upon the act, you are likely to form in him the habit of meeting the particular situation in question with total disregard. His efforts having failed so often may be so inhibited that you could hardly expect any tuition to give them new life. The matter is worse if you add further enough trials to assure you that your attracting his attention to it has been unavailing. On the other hand, if you take failure in five or ten minutes to mean inability, and from subsequent success after imitation argue that imitation was efficient, you have to face the numerous cases where animals which have failed in ten minutes have succeeded in later unaided trials. With dogs and cats this does not much matter, because they are steady performers, and their conduct in one short trial tells you what to expect with some probability. But the monkeys are much more variable and are so frequently distracted that one feels much less confidence in his predictions. Moreover, you cannot be at all sure of having attracted a monkey's attention to an object unless he does touch it. Suppose, for example, a monkey has failed to even touch a bar though you have put a bit of food on it repeatedly. It is quite possible that he may look at and take the food and not notice the bar, and the fact that after such tuition he still fails to push or pull the bar may mean simply that it has not caught his notice. I have, therefore, preferred in most cases to give the animals only a brief period of trial to test their ability by their own unaided efforts and to omit the attempts to test the efficacy of attracting their attention to the vital point in the mechanism. [p. 215]
This makes the results appear less elegant and definitive but really increases their value for purposes of interpretation.
The thoughtful reader will not expect from my experiments any perfectly rigorous demonstration of either the presence or the absence of imitation of human acts as a means of learning. The general trend of the evidence, it seems to me, is decidedly towards justifying the hypothesis that the monkeys did not learn acts from seeing me do them.
I will first describe a sample experiment and then present a summary of all those made.
On January 12th I put box Epsilon (push down) in No. 3's cage, the door of the box being open. I put a bit of food in the box. No. 3 reached in and took it. This was repeated three times. I then put in a bit of food and closed the door. No. 3 pulled and bit the box, turned it over, fingered and bit at the hole where the lever was, but did not succeed in getting the door open. After ten minutes I took the box out. Later I took No. 3 out and let him sit on my knees (I sitting on the floor with the box in front of us). I would then put my hand out toward the box and when he was looking at it would insert my finger and depress the lever with as evident a movement as I could. The door, of course, opened, and No. 3 put his arm in and took the bit of food. I then put in another, closed the door and depressed the lever as before. No. 3 watched my hand pretty constantly, as all his experiences with me had made such watching profitable. After ten such trials he was put back in the cage and the box put in with a large piece of food in it and its door closed. No. 3 failed in five minutes and the box was taken out. He was shown fifteen times more and then left to try himself. I tried him for a couple of minutes under just the same circumstances as existed during the [p. 216] tuition, i.e. he on the floor by me, the box in front. In this trial and in a five-minute trial inside his cage he failed to open the door or to differ in any essential respect from his behavior before tuition.
No. 1 saw me do 9 different acts and No. 3, 7, which they had failed of themselves to do. [5] After from 1 to 40 chances to imitate me they still failed to operate at all II of these mechanisms. In the case of 3 out of 5 that were worked the act was not the same as that taught. No. 1, who saw me pull a nail out by taking the end of it and pulling the nail away from the box, himself put his hand round the nail and wriggled it out by pulling his hand back and forth. No. 3, who saw me pull a bolt up with my fingers, succeeded by jerking and yanking the door until he shook the bolt up. He saw me pull a hook out of an eye, but he succeeded by pulling at a bar to which it was attached. In the case of one of the two remaining acts (No. 3 with nail chute) the act was done once and never again, though ample opportunity was given and tuition continued. It could, therefore, hardly have been due to an idea instilled by the tuition. The remaining case, No. 1, with loop, must, I think, be attributed to accident, especially since No. 3 failed to profit [p. 217] by precisely the same sort of tuition with precisely the same act.
Nor is there any evidence to show that although tuition failed to cause successes where unaided effort failed, it yet caused attempts which would not otherwise have occurred. Out of fifteen cases where such might have appeared, there were only three where it is possible to claim that they did. No one of these three is a sure case. With RR (wood plug) No. 1 did seem to pull the plug more definitely after seeing me than before. With QQ (c) (nail chute) and MM (bolt at top) he may possibly have done so.
In 5 cases I tried the influence of seeing me make the movement on animals who had done the act of themselves, the aim being to see whether there would be a marked shortening of the time, a change in their way of operating the mechanism or an attempt at such change. I will give the essential facts from the general table on pages 226-229.
(a) No. 1 had succeeded in pulling in the box by the upper string in 000 (upper string box) in 2.20 and then failed in 3.00. I showed him 4 times. He failed in 10. I showed him 4 more times. He failed in 10. I showed him 4 more times. He succeeded in .20. No change in manner of actor objects attacked, though my manner was different from his.
(b) No. 1 had succeeded in QQ (a) (chute bar) in 8.00. I showed him 20 times. He failed in 10. I showed him 10 more times. He succeeded in 2.00. I showed him 10 more times. He succeeded in 50 seconds. No change in his manner of performance or in the object attacked, though my manner was different from his.
(c) No. 1 had succeeded in 3.00, .25, .07, .25, .20, .06 and .09 with QQ (b) (chute bar double) and then failed in 5.00. I showed him 10 times. He then failed in 5 twice, succeeded in 3.00, and failed in 5 again. No change in manner of per- [p. 218] formance or in the object attacked, though my manner was different from his.
(d) No. 3 had the following record in box Delta:
2.00 (pushed with head)
3.20 (pushed with head)
30 F
10 F
10 F
2.10 (pulled wire and door).
I showed him 20 times by pushing the bar to the right with my finger. He succeeded in 8.00 and 8.00 by pulling the wire and the door. No change in object attacked.
(e) No. 2 had failed twice in 5 with chute QQ (ff) (chute string wire) and succeeded once in 2.00 by a strong pull or the wire itself, not the loop. I showed him 5 times, pulling the loop off the nail. He then failed in 5. There was no change in the objects attacked.
These records show no signs of any influence of the tuition that are not more probably signs of something else. We cannot attribute the rapid decrease in time taken in (b) to the tuition until we know the time-curve for the same process without tuition.
The systematic experiments designed to detect the presence of ability to learn from human beings are thus practically unanimous against it. So, too, was the general behavior of the monkeys, though I do not consider the failure of the animals to imitate common human acts as of much importance save as a rebuke to the story-tellers and casual observers. The following facts are samples: The door of No. 1's cage was closed by an iron hoop with a slit in it through which a staple passed, the door being held by a stick of wood thrust through the staple. No. 1 saw me open the door of [p. 219] his and other cages by taking out sticks hundreds of times, but though he escaped from his cage a dozen times in other ways, he never took the stick out and to my knowledge never tried to. I myself and visitors smoked a good deal in the monkeys' presence, but a cigar or cigarette given to them was always treated like anything else.
IMITATION OF OTHER MONKEYS
It would theoretically seem far more likely that the monkeys should learn from watching each other than from watching human beings, and experimental determinations of such ability are more important than those described in the last section as contributions both to genetic psychology and to natural history. I regret that the work I have been able to do in the study of this phase of the mental life of the monkeys has been very limited and in many ways unsatisfactory.
We should expect to find the tendency to imitation more obvious in the case of young and parents than elsewhere. I have had no chance to observe such cases. We should expect closely associated animals, such as members of a common troop or animals on friendly terms, to manifest it more than others. Unfortunately, two of my monkeys, by the time I was ready to make definite experiments, were on terms of war. The other had then become so shy that I could not confidently infer inability to do a thing from actual failure to do it. He showed no evidence of learning from his mates. I have, therefore, little evidence of a quantitative objective nature to present and shall have in the end to ask the reader to take some opinions without verifiable proofs.
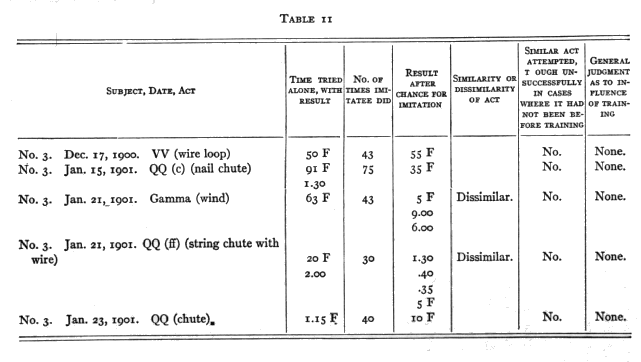
My reliable experiments, five in number, were of the following nature. A monkey who had failed of himself (and often also after a chance to learn from me or from being put [p. 220] through the act) would be put where he could see another do the act and get a reward (food) for it. He would then be given a chance to do it himself, and note would be taken of his success or failure, and of whether his act was the same as that of his model in case he succeeded, and of whether he tried that act more than before the tuition in case he tried it and failed. The results are given in Table II.
In the fourth experiment No. 1 showed further that the tuition did not cause his successes in that after some successes further tuition did not improve him.
There is clearly no evidence here of any imitation of No. 1 by No. 3. There was also apparently nothing like purposive watching on the part of No. 3. He seemed often to see No. 1 open the box or work the chute mechanism, but without special interest.
This lack of any special curiosity about the doings of their own species characterized the general behavior of all three of my monkeys and in itself lessens the probability that they learn much from one another. Nor did there appear, in the course of the three months and more the animals were together, any signs of imitation. There were indeed certain notable instances of the lack of it in circumstances which one would suppose would be favorable cases for it.
For instance: No. 2 was very timid. No. 1 was perfectly tame from the first day No. 2 was with me, and No. 3 became tame shortly after. No. 2 saw Nos. 1 and 3 come, be played with, fed and put through experiments, yet he never did the same nor did he abate a jot or tittle from his timidity save in so far as I sedulously rewarded any chance advances of his. Conversely No. 1 and No. 3 seemed uninfluenced by the fear and shyness of No. 2. No. 2's cage was between No. 1's and No. 3's, and they were for three weeks incessantly making hostile demonstrations toward [p. 221]
each other, jumping, chattering, scowling, etc. No. 2 never did anything of the sort. Again, seeing No. 3 eat meat did not lead No. 1 to take it; nor did seeing No. 1 retreat in fright from a bit of absorbent cotton lead No. 3 to avoid it.
Nothing in my experience with these animals, then, favors the hypothesis that they have any general ability to learn to do things from seeing others do them. The question is still an open one, however, and a much more extensive study of it should be made, especially of the possible influence of imitation in the case of acts already familiar either as wholes or in their elements.
LEARNING APART FROM MOTOR IMPULSES
The reader of my monograph, 'Animal Intelligence,' will recall that the experiments there reported seemed to show that the chicks, cats and dogs had only slight and sporadic, if any, ability to form associations except such as contained some actual motor impulse. They failed to form such associations between the sense-impressions and ideas of movements as would lead them to make the movements without having themselves previously in those situations given the motor impulses to the movements. They could not, for instance, learn to do a thing from having been put through it by me.
The monkeys Nos. 1 and 3 were tested in a similar way with a number of different acts. The general conclusion from the experiments, the details of which will be given presently, is that the monkeys are not proved to have the power of forming associations of ideas to any greater extent than the other mammals, that they do not demonstrably learn to do things from seeing or feeling themselves make [p. 223] the movement. An adult human being whose hand was taken and made to push in a bar or pull back a bolt would thereby learn to do it for himself. Cats and dogs would not, and the monkeys are not proved to do so. On the other hand, it is impossible for me to say, as of the dogs and cats, that the monkeys are proved not to do so. In a few cases the animals did perform acts after having been put through them which they had failed to perform when left to their own trial and success method. In the majority of cases they did not. And in some of these latter cases failure seemed so improbable in case the animal really had the power of getting an idea of the act and proceeding from idea to execution, that one is inevitably led to some explanation for the few successes other than the presence of 'ideas.'
The general manner of making these experiments was like that in the case of the cats and dogs, save that the monkey's paw was used to open the box from the outside instead of from the inside, and that the monkeys were also put through the acts necessary to operate some of the chute mechanisms. Tests parallel to that of comparing the behavior of kittens who had themselves gone into boxes with those who were dropped in by me were made in the following manner. I would carry a monkey from his cage and put him in some conspicuous place (e.g. on the top of a chair) and then give him a bit of food. This I would repeat a number of times. Then I would turn him loose in the room to see whether he had acquired an idea of being on the chair which would lead him to himself go to the chair. I would, in order to tell whether his act, in case he did so, was the result of random activities or was really due to his tuition, leave him alone for 5 or 10 minutes before the tuition. If he got on the chair afterwards when he had not before, or got on it much so would tend to show that the idea of getting food [p. 224] on that chair was present and effective. We may call these last the 'on chair' type of experiments.
A sample experiment with a box is the following: --
On January 4, 1901, box Delta (push back) was put in No. 1's cage. He failed in 5, though he was active in trying to get in for about 4 minutes of the time and pulled and pushed the bar a great deal, though up and down and out instead of back. In his aimless pushings and pullings he nearly succeeded. He failed in 5 in a second trial also. I then opened the door of the cage, sat down beside it, held out my hand, and when he came to me took his right paw and with it (he being held in front of the box) pushed the bar back (and pulled the door open in those cases when it did not fall open of itself). He reached in and took the food and went back to the top of his cage and ate it. (No. 1 generally did this, while No. 3 generally stayed by me.) I then tried him alone; result 10 F ; no activity at all. On January 5th I put the box in; result 10 F. He was fairly active. He pulled at the bar but mostly from a position on the top of the box and with his left hand; no attempts like the one I had tried to teach him. Being left alone he failed in 5. Being tried again with the door of the cage open and me sitting as I had done while putting him through the act, he succeeded in 7.00 by pushing the bar with his head in the course of efforts to poke his head in at the door. I then put him through the act 10 times and left him to himself. He failed in 5.00; no activity. I then sat down by the cage as when teaching him. He failed in 5; little activity. Later in the day I put him through the act 10 times and then left him to himself. He failed in 5; little activity. I sat down as before. He failed in five; little activity. On January 6th I put him through the act 10 times and then left him. He failed in 10. This was repeated later in the day with the same result. [p. 225]
Record: - By himself, 10 F. Put through 80 times. F 65 (a) {[the (a) refers to a note of his unrepeated chance success with his head]}. No similar act unsuccessfully attempted. Influence of tuition, none.
With the chute mechanisms the record would be of the same nature. With them I put the animal through generally by taking his paw, held out through the wire netting of the cage, and making the movement with it. In one experiment (No. 3 with QQ chute) the first 58 trials were made by taking the monkey outside the cage and holding him instead of having him put his paw through the netting for me to take.
Many of the experiments were with mechanisms which had previously been used in experiments concerning the ability to learn from seeing me operate them. And the following Table (12) includes the results of experiments of both sorts. The results of experiments of the ` on chair' type are in Table 13. In cases where the same apparatus was used for both purposes, the sort of training which was given first is that where an A is placed.
In the first four experiments with No. 1 there was some struggling and agitation on his part while being held and put through the act. After that there was none in his case except occasional playfulness, and there was never any with No. 3 after the first third of the first experiment. The monkeys soon formed the habit of keeping still, because it was only when still that I put them through the act and that food resulted. After you once get them so that they can be held and their arms taken without their clinging to you, they quickly learn to adapt themselves to the experiments.
With No. 1, out of 8 cases where he had of himself failed (in five of the cases he had also failed after being shown by me), he succeeded after being put through (13, 21, 51, 10, 7, [p. 226]
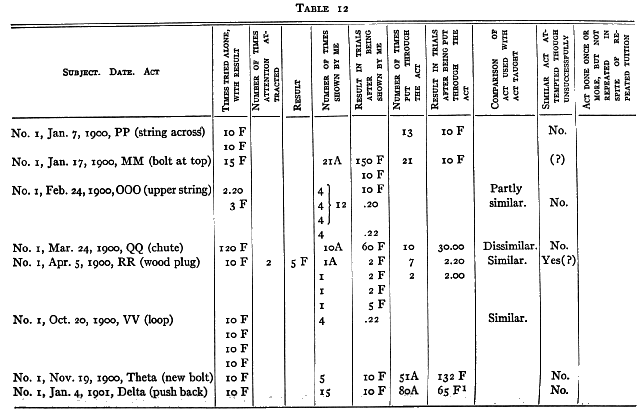
[p. 227]
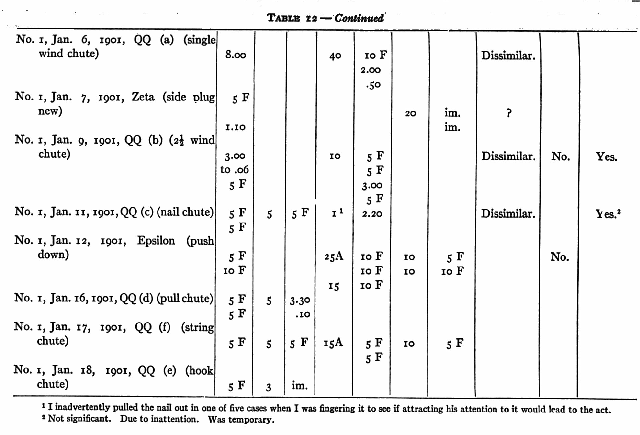
[p. 228]
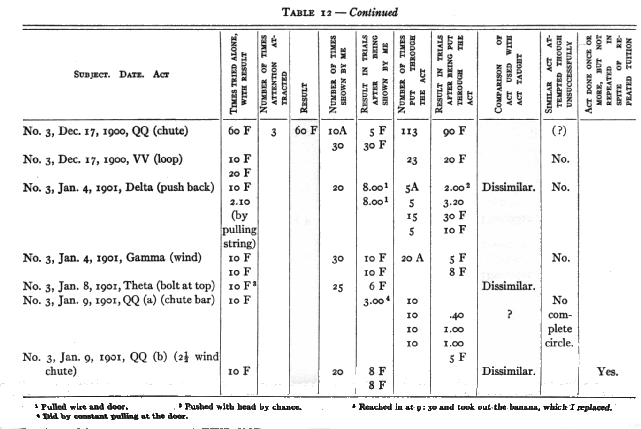
[p. 229]
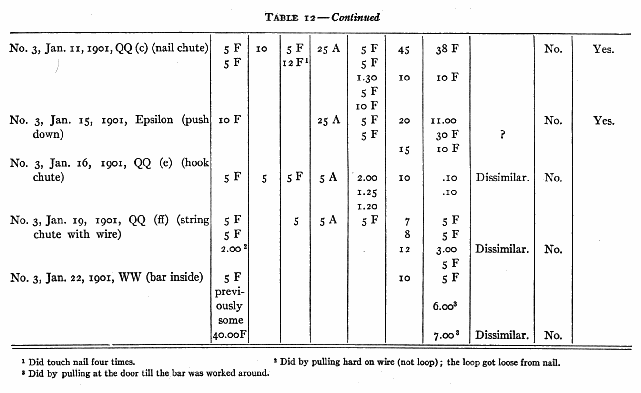
[p. 230] 80, and 10 times) in two cases (QQ (chute) and RR (wood plug). The act was unlike the one taught him in the former case.
In only one case (bolt at top) out of eight was there possibly any attempt at the act after he had been put through which had not been made before. The 'yes or ?' in the table with RR was a case occurring after the imitation of me but before the putting No. 1 through.
Out of 6 cases where he had himself failed, No. 3 succeeded (after being put through 113, 23, 20, 10, 10, 20 and 10 times) in 3 cases (chute bar, push down and bar inside). The act was dissimilar in all three cases, bearing absolutely no resemblance in one case. There was no unsuccessful attempt at the act taught him in any of the cases. With the chute he did finger the bar after tuition where he had not done so before, but it was probably an accidental result of his holding his hand out toward it for me to take as he had formed the habit of doing. In the case of box Epsilon (push down), with which he succeeded by pushing his hand in above the lever (an act which though unlike that taught him might be by some considered to be due to an idea gained from the tuition), he failed entirely after further tuition (15 times).
Like the dogs and cats, then, the monkeys seemed unable to learn to do things from being put through them. We may now examine those which they did do of themselves before tuition and ask whether they learned the more rapidly thereby or modified their behavior in ways which might be due to the tuition. There are too few cases and no chance for comparison on the first point; on the second the records are unanimous in showing no change in the method of operating the mechanisms due to the tuition.
As in Table 9, figures followed by F mean that in that [p. 231] length of time the animal failed. Figures without an F denote the time taken by the animal to operate the mechanism.
As a supplement to Table 12 I have made a summary of the cases where the animals did succeed after tuition, that shows the nature of the act shown them as compared with the act they made use of.
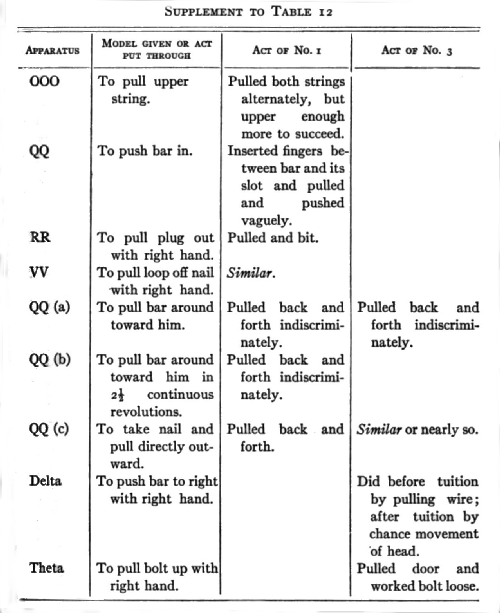
[p. 232]
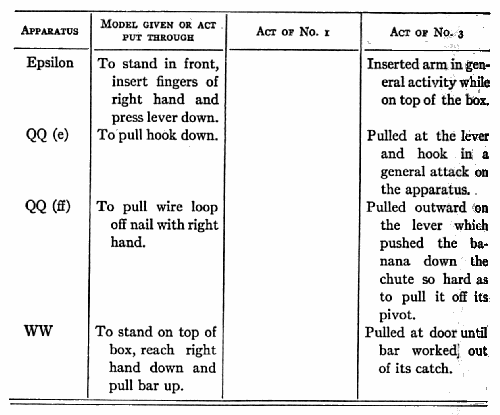
I have kept the results of the tests of the 'on chair' type separate from the others because they may be tests of a different thing and surely are subject to different conditions.
They were tests of the animals' ability to form the habit of going to a certain place by reason of having been carried there and securing food thereby. I would leave the animal loose in the room, and if he failed in 5 or 10 minutes to go to the place of his own accord, would put him back in his cage; if he did go of his own accord, I would note the time. Then I would take him, carry him to the place, and feed him. After doing this 10 times I would turn him loose again and see whether the idea of being fed in such and such a place was present and active in making him go to the place. In such tests we are absolutely sure that the animal can without any difficulty perform the necessary movements and would in [p. 233] case the proper stimulus to set them off appeared, if, for instance, a bit of food on one of the places to which he was to go caught his eye. In so far forth the tests were favorable cases for learning. On the other hand, the situation associated with getting food may have been in these cases not the mere 'being on box' but the whole previous experience 'being carried while clinging and being put or let jump on a box.' In this respect the tests may have been less favorable than the acts where getting food was always the direct sequent of the act of going into the box.
The experiments were: -
A. Carrying the animal and putting him on a chair.
B. Carrying the animal and putting him on a pile of boxes.
C. Carrying the animal and putting him on the top of a sewing machine.
D. Carrying the animal and putting him on the middle of a board 6 feet long, stretched horizontally across the room, 3 feet from the floor.
E. Carrying the animal and putting him on the side of the cage, head down.
The results are given in Table 13.
The size of the room in which I worked and other practical difficulties prevented me from extending these experiments. As they stand, no stable judgments can be inferred from them. It should be noted that in the successful cases there were no other signs of the presence of the idea 'food when there' than the mere going to a certain place. The animal did not wait at the place more than a second or two, did not look at me or show any signs of expecting anything.
Although, as I noted in the early part of this monograph, there were occasionally phenomena in the general behavior of the monkeys which of themselves impressed one as being suggestive of an ideational life, the general run of their [p. 234]
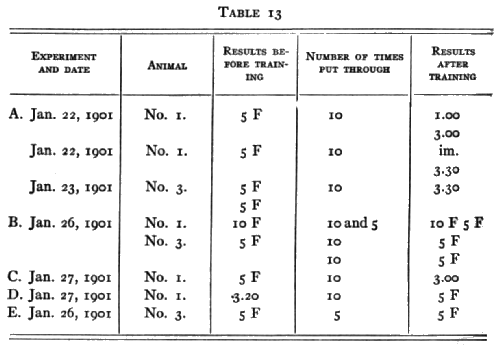
learning apart from the specific experiments described was certainly confined to the association of impulses of their own with certain situations. The following examples will suffice: -
In getting them so that they would let themselves be handled it was of almost no service to take them and feed them while holding them or otherwise make that state pleasant for them. By far the best way is to wait patiently till they do come near, then feed them; wait patiently till they do take hold of your arm, then feed them. If you do take them and hold them partly by force, you must feed them only when they are comparatively still. In short, in taming them one comes unconsciously to adopt the method of rewarding certain of their impulses rather than certain conditions which might be associated in their minds with ideas, had they such.
After No. 1 and No. 3 had both reached a point where both could hardly be gotten to leave me and go back into [p. 235] their cages or down to the floor of the room, where they evidently enjoyed being held by me, they still did not climb upon me. The idea of clinging to me was either absent or impotent to cause them to act. What they did do was, in the case of No. i, to jump about, pawing around in the air, until I caught an arm or leg, to which stimulus he had by dint of the typical sort of animal association learned to react by jumping to my arm and clinging there; in the case of No. 3, to stand still until I held my arm right in front of him (if he were in his cage) or to come and stand on his hind legs in front of me (if he were out on the floor). In both cases No. 3's act was one which had been learned by my rewarding his impulses. I often tried, at this period of their intimacy with me, this instructive experiment. The monkey would be clinging to me so that I could hardly tear him away. I would do so, and he would, if dropped loose from me, make no efforts to get back.
I have already mentioned my failure to get the animals to put out their right hands through the netting after they had long done so with their left hands. With No. 3 I tried putting my fingers through and poking the arm out and then making the movement with it. He profited little if any by this tuition. Had I somehow induced him to do it himself, a few trials would have been sufficient to get the habit well under way.
Monkey No. 1 apparently enjoyed scratching himself. Among the stimuli which served to set off this act of scratching was the irritation from tobacco smoke. If any one would blow smoke in No. 1's face, he would blink his eyes and scratch himself, principally in the back. After a time he got in the habit of coming to the front of his cage when any one was smoking and making such movements and sounds as in his experience had attracted attention and [p. 236] caused the smoker to blow in his face. He was often given a lighted cigar or cigarette to test him for imitation. He formed the habit of rubbing it on his back. After doing so he would scratch himself with great vigor and zest. He came to do this always when the proper object was given him. I have recounted all this to show that the monkey enjoyed scratching himself. Yet he apparently never scratched himself except in response to some sensory stimulus. He was apparently incapable of thinking 'scratch' and so doing. Yet the act was quite capable of association with circumstances with which as a matter of hereditary organization it had no connection. For by taking a certain well defined position in front of his cage and feeding him whenever he did scratch himself I got him to always scratch within a few seconds after I took that position.
GENERAL MENTAL DEVELOPMENT OF THE MONKEYS
It is to be hoped that the growing recognition of the worth of comparative and genetic studies will lead to investigations of the mental make-up of other species of monkeys, and to the careful overhauling of the work done so far, including these rather fragmentary studies of mine. Work with three monkeys of one species, especially when no general body of phenomena, such as one has at hand in the case of domestic animals, can be used as a means of comparison, must necessarily be of limited application in all its details and of insecure application even in its general features. What I shall say concerning the advance in the mental development of the monkeys over that of other mammals may then be in strictness true of only my three subjects, and it may be left to the judgment of individuals to extend my conclusions [p. 237] as far as seems to them likely. To me it seems fairly likely that the very general mental traits which the research has demonstrated hold true with little variation in the monkeys in general.
The monkeys represent progress in mental development from the generalized mammalian type toward man: -
1. In their sensory equipment, in the presence of focalized vision.
2. In their motor equipment, in the coordinated movements of the hand and the eye.
3. In their instincts or inherited nervous connections, in their general physical and mental activity.
4. In their method of learning or associative processes; in-
a. Quicker formation of associations,
b. Greater number of associations,
c. Greater delicacy of associations,
d. Greater complexity of associations,
e. Greater permanence of associations.
The fact of (1) is well known to comparative anatomists. Its importance in mental development is perhaps not realized, but appears constantly to a systematic student.
(2) is what accounts for much of the specious appearance of human ways of thinking in the monkeys and becomes in its human extension the handy tool for much of our intellectual life. It is in great measure the prerequisite of 4. c.
(3) accounts for the rest of such specious appearances, is at the basis of much of 4. b, presages the similar though extended instincts of the human being, which I believe are the leading efficient causes of human mental capacity, and is thus the great mental bond which would justify the inclusion of monkeys and man in a common group if we were to classify animals on the basis of mental characteristics. [p. 238]
Even the casual observer, if he has any psychological insight, will be struck by the general, aimless, intrinsically valuable (to the animal's feelings) physical activities of a monkey compared with the specialized, definitely aroused, utilitarian activities of a dog or cat. Watch the latter and he does but few things, does them in response to obvious sense presentations, does them with practical consequences of food, sex-indulgence, preparation for adult battles, etc. If nothing that appeals to his special organization comes up, he does nothing. Watch a monkey and you cannot enumerate the things he does, cannot discover the stimuli to which he reacts, cannot conceive the raison détré of his pursuits. Everything appeals to him. He likes to be active for the sake of activity.
The observer who has proper opportunities and takes proper pains will find this intrinsic interest to hold of mental activity as well. No. 1 happened to hit a projecting wire so as to make it vibrate. He repeated this act hundreds of times in the few days following. He did not, could not, eat, make love to, or get preliminary practice for the serious battles of life out of, that sound. But it did give him mental food, mental exercise. Monkeys seem to enjoy strange places; they revel, if I may be permitted an anthropomorphism, in novel objects. They like to have feelings as they do to make movements. The fact of mental life is to them its own reward.
It is beyond question rash for any one to venture hypotheses concerning the brain parallel of mental conditions, most of all for the ignoramus in the comparative histology of the nervous system, but one cannot help thinking that the behavior of the monkeys points to a cerebrum that is no longer a conservative machine for making a few well-defined sorts of connections between sense impressions and acts, [p. 239]
that is not only fitted to do more delicate work in parts, but is also alive, tender all over, functioning throughout, set off in action by anything and everything. And if one adds coördinations allowing a freedom and a differentiation of action of the muscles used in speech comparable to that already present in connection with the monkey's-hand, he may well ask, "What more of a nervous mechanism do you need to parallel the behavior of the year-old child?" However, this is not the place to speculate upon the importance to human development of our instinctive aimless activity, physical and mental, or to describe further its similarity and evident phylogenetic relationship to the instinctive behavior of the monkeys. Elsewhere I shall undertake that task.
4. In their method of learning, the monkeys do not advance far beyond the generalized mammalian type, but in their proficiency in that method they do. They seem at least to form associations very much faster, and they form very many more. They also seem superior in the delicacy and in the complexity of the associations formed and the connections seem to be more permanent.
This progress may seem, and doubtless will to the thinker who looks upon the human intellect as a collection of functions of which ideation, judgment and reasoning are chief, to be slight. To my mind it is not so in reality. For it seems to me highly probable that the so-called 'higher' intellectual processes of human beings are but secondary results of the general function of having free ideas and that this general function is the result of the formation after the fashion of the animals of a very great number of associations. I should therefore say, "Let us not wonder at the comparative absence of free ideas in the monkeys, much less at the absence of inferences or concepts. Let us not wonder [p. 240] that the only demonstrable intellectual advance of the monkeys over the mammals in general is the change from a few, narrowly confined, practical associations to a multitude of all sorts, for that may turn out to be at the bottom the only demonstrable advance of man, an advance which in connection with a brain acting with increased delicacy and irritability, brings in its train the functions which mark off human mental faculty from that of all other animals.
The typical process of association described in Chapter II has since been found to exist among reptiles (by Mr. R. M. Yerkes) and among fishes (by myself). It seems fairly likely that not much more characterizes the primates. If such work as that of Lubbock and the Peckhams holds its own against the critical studies of Bethe, this same process exists in the insects. Yerkes and Bosworth think they have demonstrated its presence in the crayfish. Even if we regard the learning of the invertebrates as problematic, still this process is the most comprehensive and important thing in mental life. I have already hinted that we ought to turn our views of human psychology upside down and study what is now casually referred to in a chapter on habit or on the development of the will, as the general psychological law, of which the commonly named processes are derivatives. When this is done, we shall not only relieve human mentality from its isolation and see its real relationships with other forms; we may also come to know more about it, may even elevate our psychologies to the explanatory level and connect mental processes with nervous activities without arousing a sneer from the logician or a grin from the neurologist.
Footnotes
[1] This chapter appeared originally as Monograph Supplement No. 15 to the Psychological Review.
[2] Pp. 20 to 155 of this volume.
[3] American Journal of Psychology, Vol. X, Pp. 256-279; Vol. X, Pp. 80-100; Vol. XI, Pp. 206-239.
[4] This, I regret, was not done [E. L. T., 1911].