Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
TWO TYPES OF CONDITIONED REFLEX AND A PSEUDO TYPE[*]
From the Biological Laboratories of Harvard University
B.F. Skinner (1935)[1]
First published in Journal of General Psychology, 12, 66-77.
A conditioned reflex is said to be conditioned in the sense of being
dependent for its existence or state upon the occurrence of a certain kind
of event, having to do with the presentation of a reinforcing stimulus.
A definition that includes much more than this simple notion will probably
not be applicable to all cases. At almost any significant level of analysis
a distinction must be made between at least two major types of conditioned
reflex. These may be represented, with examples, in the following way (where
S = stimulus, R = response, (S - R) = reflex, ->
= "is followed by," and [ ] = "the strength of" the inclosed reflex):
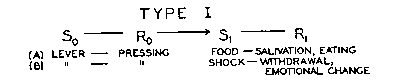
Given such a sequence, where [S1-R1]
~ = 0,[2] conditioning occurs as a change in [So-Ro]
- an increase in strength (positive conditioning) in (a) and a decrease
(negative conditioning) in (b).[p.67]
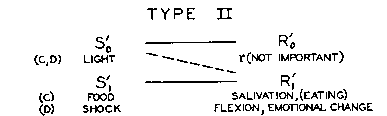
Given the simultaneous or successive presentation of S'o and
S'1, where [S'1- R'1]
~ =0,[2] conditioning occurs as an increase in [S'o
- R'1].
Differences between the types are as follows:
1. In Type I, So ->Ro->
S1, where Ro necessarily intervenes
between the stimuli; in Type II, S'o ->S'1,
where R'o is ignored.[3] In I, Ro
is important; it becomes the conditioned response. In II, R'o
is irrelevant (except when it is relevant in another sense by conflicting
with R'1) and may actually disappear.
Since conditioning of the second type may take place even when S'1
occurs after R'o, Paradigm II, Example (c),
may be written for this case as follows:
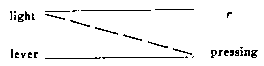
light --------------- r --------------->
food ---------------salivation,
when it is identical with I. But the result is not to reduce the two
types to a single form. Both kinds of conditioning proceed simultaneously
but separately. If r is "turning toward the light," for example,
and if the food is withheld until turning takes place, [light - turning]
will increase according to I while [light-salivation] will increase according
to II. The same result is obtained with negative conditioning. Example
(d) gives, upon delaying S'1,
light --------------- r --------------->
shock --------------- flexion,
etc.,
where [light--r] will decrease according to I, while [light-flexion]
increases according to II.
In the special case in which Ro and R'1
are of the same form, the two kinds can apparently not be separated. If,
for example, some unconditioned salivation is supposed to be elicitable
by a light,[4] we may substitute "salivation" for r,
to obtain
light - salivation I --------------->
food - salivation II.
Both [light - salivation I] and [light - salivation II] will increase,
with apparently no possible distinction. This is a very special case and
is also in no sense a reduction to a single type.
2. In I, (So - Ro) occurs normally
in the absence of (S1 - R1), [p.68]
and its strength may be measured without interfering with the reinforcing
action of S1. In II S'1 must be withheld
whenever a measurement of [S'o - R'1],
the conditioned reflex, is taken, because S'1 also evokes
R'1. Some amount of extinction necessarily ensues in
the second case.
3. Since S'1 must be withheld in measuring [S'o
- R'1], R'1 must be independent
of any property of S'1 not possessed by S'o.
In Example (c) salivation may become attached to the light as a
conditioned response of Type II; but seizing, chewing, and swallowing,
which are also responses to S'1, must not be included
in the paradigm since they require the presence of parts of S'1
which cannot be supplied by S'o.
A special restriction on Paradigm II is therefore necessary. Where S'o
is of a very simple sort (a tone, for example), the properties possessed
in addition to S'o by S'1 are practically
equal to S'1, and we may express the restriction in terms
of a general distinction between two kinds of response. The first kind
require no external point of reference in their elicitation or description.
Typical examples are: glandular activities (salivation), local muscular
responses (flexion, wink, breathing movements, production of sounds), and
facilitation and inhibition.[5]
The second kind require points of reference for their elicitation or
description which are not supplied by the organism itself, but by the stimulus.
Examples are: orientation toward the source of a sound, approaching a light,
and touching, seizing and manipulating objects (such as a lever or food).
Our present rule is that responses of the second kind cannot be substituted
for R'1 in Paradigm II, unless S' also supplies
the required points of reference.
4. In Type I, [So - Ro] ~ = 0 before
conditioning takes place. The reflex-to-be-conditioned must be elicited
at least once as an unconditioned "investigatory" reflex. In Type II, [S'o
- R'1] may begin at zero and usually does. In Type
I the state of the reflex is "conditioned" by the occurrence of
the reinforcing sequence, but its existence is not. A distinction
between a conditioned and an unconditioned relfex is here less significant,
because all examples of the [p.69] former have necessarily been examples
of the latter. There are no exclusively conditioned reflexes in this type.
Since [S'o - R'1] may begin at zero,
a new reflex may be created in conditioning of the second type. And since
practically any stimulus may be attached to R'1 in Paradigm
II, a very large number of new reflexes can thus be derived. Conditioning
of Type I, on the other hand, is not a device for increasing the repertory
of reflexes; Ro continues to be elicited by the one stimulus
with which it began.
There are three reflexes in Paradigm II, but only two in I.
5. The significant change in Type I may be either an increase or a decrease
in strength; in Type II it is an increase only, even when [S'o
- R'1] does not begin at zero.
In Type I stimuli may be divided into two classes, positively and negatively
conditioning, according to whether they produce an increase or decrease
when used as reinforcement. The distinction cannot be made in Type II,
where a reflex may be negative in another sense (a reflex of "avoidance,"
for example), but where its strength only increases during conditioning.
6. In Type I the conditioned reflex (So - Ro)
may be associated with any drive; in Type II the reflex (S'o -
R'1) is necessarily attached to the drive specified
by R'1.
This point may require some comment. In the present use of the term
a drive is an inferred variable of which the strength of a group of reflexes
is a function (2). Hunger, for example, is a variable (H) a change
in which is responsible for concurrent changes in the strength (a)
of all unconditioned reflexes concerned with the ingestion of food, (b)
of all conditioned reflexes (of either type) in which the reinforcing stimulus
is concerned with the ingestion of food, and (c) to a much lesser
extent of all "investigatory" reflexes. In Paradigm I, Example a
(lever - pressing) is originally a function of H to some slight
extent under (c) above. After conditioning it varies with H according
to (b), over a wide range probably equal to that of any unconditioned
reflex under (a). Conditioning of Type I is really the becoming
attached to a group of reflexes varying as a function of some drive. This
is a much more comprehensive description of the process than to define
it as an increase in strength, where the drive is assumed to remain constant
at a significant value. But the identity of H in the present case
is [p.70] determined only by our choice of a reinforcing reflex. Given
(S1 - R1) of another drive, say thirst,
then (So - Ro) will become conditioned
by attaching itself to the group varying with thirst, and will not vary
with H except to some slight extent under (c).
This is a characteristic wholly lacking in Type II. Here R' is
originally part of the unconditioned reflex and the drive to which it belongs
is definitely fixed.
7. A minor difference is in the way in which the stimulus-to-be-conditioned
usually acts. In Type I, So is usually part of a larger
field, and Ro occurs as the result of the eventual prepotency
of So over other stimuli. In Type II, S'o
is usually suddenly presented to the organism. The significance of
this difference, which is not absolute, will appear later.
We shall now consider a third type of relation which involves a discrimination.
It may be based upon a conditioned reflex of either type, but we shall
begin with I. To establish a discrimination subdivide So into
two classes on the basis of a selected property or component member (3,
4). For example, let the lever stimulate either in the presence of
a light (L), when the stimulus may be written as SA B..L..(subcripts
indicate properties or components), or in the dark, when the stimulus is
SA.B.... Continue to reinforce the re-response to one
of them, say SA.B..L.., and extinguish or negatively
condition the response to the other by breaking the sequence at S1
or by introducing as S1 of the negatively conditioning
kind (Difference 5). When this has been done, [SA.B..L.---Ro]
> [SA.B..---Ro]. And at any value of
the underlying drive such that (SA.B..L ---Ro)
is usually elicited but (SAB ---Ro)
is not, there exists the following condition. Given an organism in the
presence of SAB.. ordinarily unresponsive, the presentation
of L will be followed by a response. For the sake of comparison
we may set up a paradigm in imitation of II as follows:
The relation between the light and the response to the lever might
be called pseudo-conditioned reflex. It has some of the characteristics
of Type II: the original response to the light is irrelevant [p.71] (Difference
1); the relation may be wholly absent prior to the "conditioning" (Difference
4); it changes in a positive direction only (Difference 5); and the "stimulus"
is usually of the presented kind (Difference 7). In all these respects
it differs from Type I, although the example is based upon a reflex of
that type. I many other respects it differs from both types. A reinforcing
reflex is not included in the paradigm, but must be added as a third or
fourth reflex. The response is not principally to the light, but to the
lever; the light is only a component member of the whole stimulus, and
"light-pressing" is not legitimately the expression of a reflex. The lever
cannot be removed to show the conditioned effectivelness of the light as
in Type II; instead, the response to the lever alone must be extinguished
- a characteristic that we have not met before.
In spite of these differences it is often said (in similar cases) that
the light becomes the "conditioned stimulus for the response to the lever"
just as it becomes the stimulus for salivation. This is a confusion with
Type II which obviously arises from a neglect of the extinguished reflex.
The relation of pressing the lever to the lever itself is ignored and only
the relation to the light taken into account. The lever comes to be treated,
not as a source of stimulation, but as part of the apparatus, relevant
to the response only for mechanical reasons. When the discrimination is
based upon a response not requiring an external point of reference (Difference
3), the chance of this neglect increases enormously. If we substitute "flexion
of a leg" for "pressing a lever" (and continue for the moment with Type
I), So in Paradigm I is not directly observable; we simply
wait until a flexion appears, then reinforce. Having established (So
- Ro) as a conditioned reflex of some strength,
we subdivide our inferred So as before, extinguish (SAB..-
Ro), and reinforce (SAB..L..- Ro).
When the discrimination has been set up, we have a condition in which the
organism is ordinarily unresponsive but immediately responds with flexion
upon presentation of the light.
Our inability to demonstrate So makes it difficult
to show the discriminative nature of this relation; but it is by no means
impossible to find other grounds, as we may see by comparing it with a
true reflex of Type II. Let the presentation of the light be followed by
a shock to the foot until the light alone elicits flexion. The resulting
reflex is superficially similar to the relation of light and [p.72] flexion
that we have just examined, but fundamentally the two cases are unlike.
Assuming that no immediate difference can in fact be detected,[6]
we may still show differences by referring forward or backward to the history
of the organism. The two relations have been established in different ways
and their continued existence depends upon reinforcement from different
stimuli. The discriminative drive relation also varies with an arbitrarily
chosen drive, while the conditioned reflex is necessarily attached to the
drive to which shock-flexion belongs.
These differences are chiefly due, however, to the use of a conditioned
reflex of Type I in setting up the discrimination. In a pseudo-conditioned
reflex based upon Type II the distinction is much less sure. Here we are
invariably able to neglect the extinguished member because R'1
is of the kind not requiring an external point of reference (Difference
3), and we can minimize its importance in other ways. Given a conditioned
reflex of this kind:
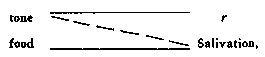
if we establish a discrimination between the tone and the tone-plus-a
light (reinforcing the resonse to the latter), we obtain the following
condition: an organism in the presence of the tone, ordinarily unresponsive,
will respond upon presentation of the light. The only difference between
this relation and a true reflex of Type II is the extinction of the response
to the tone, which is evidence that a discrimination has taken place. The
reinforcement of tone and light should condition responses to both of these
stimuli; but we observe that the organism is unresponsive in the presence
of the tone alone.
Now, this surviving difference may be reduced at will by reducing the
significance of S'o in the basic reflex of the pseudo-type.
If we lower the intensity of the tone or choose another stimulus of a less
important kind, we may approach as closely as we please to a conditioned
reflex of Type II. We cannot actually reach Type II in [p.73] this way,
but we can easily reach a point at which our pseudo-reflex is identical
with any actual experimental example of that type. This is true because
some amount of discrimination is practically always involved in cases of
Type II. When we put a dog into a stand, present a light and then food,
the food reinforces not only the light but the stimulation from the stand.
Merely putting the dog into the stand again should elicit salivation according
to Paradigm II. In practice this is a disturbing effect, which must be
eliminated through extinction. So long as it occurs, any actual case of
Type II must be formulated as a pseudo-conditioned reflex. If SG
is the stimulation affecting the organism in addition to So,
then So Paradigm II should read SG +
So. The effect upon SG is extinguished
through lack of reinforcement in the absence of So, and
the result is a discrimination: an organism is the presence of SG,
ordinarily unresponsive, responds when So is added. The
importance of this criticism will depend upon the relative magnitudes of
SG and So. In the optimal experiment
SG may be reduced to a value that is insignificant in
comparison with ordinary values of So.
The partially discriminative nature of Type II is invevitable. It is
not important in Type I because of Difference 1. Paradigm I contains an
implicit specification that So is active or has just
acted at the moment of reinforcement, since it specifies that S1
is to be withheld until Ro has occurred. The reinforced
stimulus is really So and not SG +
So (it is the lever, in our example, not the whole stimulating
field presented by the apparatus). Paradigm II contains no specification
of the activity of So ; and the reinforcing action of
S'1 must be supposed to extend to SG as
well as to So. In practice an active state at the moment
of reinforcement is usually insured by presenting So suddenly.[7]
This might be included as an additional provision in Paradigm II, but the
provision really required is that So, and no part of
SG, be active at the moment of reinforcement. This is
not easily arranged. We cannot wholly avoid the generalized action of the
reinforcement in Type II because of the lack of dependence of S'1
upon R'o.
One characteristic of the pseudo-conditioned reflex is the variety [p.74]
of the forms of its "stimulus". We have assumed that in our two fundamental
paradigms any stimulus had ultimately the dimensions of energy (although
we have often used the shorthand device of speaking of the source of the
energy - as, for example, "lever"). In the pseudo type, however, the "stimulus"
can be a single property. It can be the intensity of the stimulus, or some
such qualitative aspect as pitch or hue. It can be a change from one value
of a property to another, or the absence of a property, or a duration.
The reason why this is possible is that the other properties of the stimulus
can be relegated to SG for extinction. If the pitch
of a tone is to be a conditioned "stimulus," the tone itself must first
become one also, and the response to its other properties must be extinguished
by extinguishing the responses to tones of other pitches. In a true conditioned
reflex this cannot be done. Although it is common to speak of properties
as stimuli (1), the presence of a property in the position of a stimulus
is a certain indication that a pseudo-conditioned reflex is really in quesiton.
A property alone cannot be used in either true type because it implies
extinction; most of the real stimulus must be relegated to SG,
and the requirement that the value of SG be negligible
cannot therefore be satisfied.
The position of a pseudo-conditioned reflex may be summarized as follows.
When the pseudo-reflex is based upon a reflex of Type I and when Ro
requires external points of reference, there are important practical
and theoretical reasons why a separate formulation is demanded. When Ro
does not require external points of reference, there are fewer
differences, but a separate formulation is still necessary. When the pseudo
type is based upon a reflex of Type II, the distinction is weakened but
should still be made, except when SG can be reduced to
a very low value relative to S'o. In the last case a
practical distinction is impossible, not because of an identity of types,
but because of the failure of Type II to appear experimentally in a pure
form.
It is a tempting hypothesis that II is not an authentic type but may
be reduced to a discrimination based on Type I. But this has not been shown;
we have not reduced the pseudo type to Type II or vice versa. Nor
have we come very near it. The present pseudo-reflex which resembles II
most closely requires of that type for its establishment. It is probably
more than a coincidence that [p.75] a discrimination based upon Type I
has so many of the properties of II, but the reduction to a single type
appears from our present evidence to be highly improbable, desirable though
it would be as an immense simplification. The differences that we have
noted are not easily disposed of. Still more improbable is a reduction
of I to II, since the first step supplied by the pseudo type is then lacking.
To the differences we have listed might be added differences in the
parts played by the two types in the economy of the organism. The essence
of Type II is the substitution of one stimulus for another, or, as Pavlov
has put it, signalization (1). It prepares the organism by obtaining
the elicitation of a response before the original stimulus has begun to
act, and it does this by letting any stimulus that has incidently accompanied
or anticipated the original stimulus act in its stead. In Type I there
is no substitution of stimuli and consequently no signalization. Type I
acts in another way: the organism selects from a large repertory of unconditioned
reflexes those of which the repetition is important with respect to certain
elementary functions and discards those of which it is unimportant. The
conditioned response of Type I does not prepare for the reinforcing stimulus,
it produces it. The stimulus-to-be-conditioned is never in any sense
incidental.
Type I plays the more important rôle. When an organism comes accidentally[8]
upon a new kind of food, which it seizes and eats, both kinds of conditioning
presumably occur. When the visible radiation from the food next stimulates
the organism, salivation is evoked according to Paradigm II. This secretion
remains useless until the food is actually seized and eaten. But seizing
and eating will depend upon the same accidental factors as before unless
conditioning of Type I has also occurred - that is, unless the strength
of the reflex (food-seizing) has increased. Thus while a reflex of Type
II prepares the organism, a reflex of Type I obtains the food for which
the preparation is made. And this is in general a fair characterization
of the relative importance of the two types. As Pavlov has said, conditioned
stimuli are important in providing saliva before food is received, but
"even greater is their importance when they evoke the motor component of
the complex reflex of nutrition, i.e., [p.7 6] when they act as stimuli
to the reflex of seeking food." (1, p.13).[9] Although "the reflex of seeking
food" is an unfortunate expression, it refers clearly enough to behavior
characteristic of Type I.
Footnotes
[*] Accepted for publication by Carl Murchison of the
Editorial Board and received in the Editorial Office, June 4, 1934.
[1] Society of Fellows, Harvard University.
[2] This expression specifies the presence of some
amount of drive (2).
[3] For convenience we shall omit the case of simultaneous
stimuli in Type II.
[4] See Difference 4 below for this general requirement
in Type I.
[5] Where conditioned facilitation and inhibition are
defined by substituting for R'1 in Paradigm II the expressions
"Incr.
[S" - R"]" and "Decr. [S" - R"]" respectively.
(Incr. = "increase in"; Decr. = "decrease in").
[6] This is a generous assumption since some evidence
for the presence of So can usually be found. A difference
in the character of the response might also be shown (in the case of the
true reflex it may be accompanied by changes in breathing rate, for example,
which would be lacking in the pseudo reflex).
[7] This is our explanation of Difference 7. Another
explanation might be added. If S'o is active for any
length of time prior to S1 it will have an extinguishing
effect. This cannot be said of Type I.
[8] That is to say, as the result of weak investigatory
reflexes.
[9] This is a doubly interesting statement because
Pavlov has confined his own investigations practically exclusively to conditioned
reflexes of the second type. It ought to be said that he usually regards
this type as adequate for the whole field. Thus he says that the "function
of the hemispheres" is signalization (1, p.17), although signalization
is, as we have seen, a characteristic of Type II only.
References
1. Pavlov, I.P. Conditioned reflexes (Trans. & ed. by G.V. Anrep)
London: Oxford Univ. Press, 1927. Pp xvi + 430.
2. Skinner, B.F. Drive and reflex strength: I. J. Gen. Psychol.,
1932, 6, 22-37.
3. ----------. The rate of establishment of a discrimination. J.
Gen. Psychol., 1933, 9, 302-350.
4. ----------. The generic nature of the concepts of stimulus and response.
J. Gen. Psychol., 1935, 12, 40-65.
Harvard University
Cambridge, Massachusetts
DEUX TYPES DU RÉFLEXE CONDITIONEL ET UN PSEUDO-TYPE
(Résumé)
On définit deux types principaux du réflexe conditionnel
et donne une liste des différences entre eux. Un pseudo-réflexe
conditionel, où il s'agit d'une discrimination, peut être
basé sur le Type I pour donner quelques-unes des propriétés
du Type II. Quand it est basé sur le Type II il se peut dans un
cas spécial, qu'on ne puisse le distinguer de ce type, mais cela
est dû au manque du Type II de se montrer expérimentalement
dans une forme pure et n'indique pas une réduction à un seul
type. Dans une pseudo-réflexe le "stimulus" peut avoir la forme
d'une seule propriété, mais cela n'est possible dans l'un
ni l'autre vrai type. On note la place de chaque type dans l'économie
de l'organisme.
Skinner
ZWEI TYPEN DES BEDINGTEN REFLEXES UND EIN PSEUDOTYPUS
(Referat)
Zwei Haupttypen des bedingten Reflexes werden bestimmt und Unterschiede
zwischen ihnen angegeben. Ein pseudobedingter Reflex, der eine Unterscheidung
voraussetzt kann auf Typus I beruhen, so dass er einige der Eigenschaften
von Typus II aufweist. Wenn er auf Typus II beruht, kann er in einem besonderem
Fall nicht unterscheidbar von jenem Typus sein, aber dies ist die Folge
der Nichterscheinung des Typus II experimentell in einer reinen Form und
beweist eine Zurückführung auf einen einzelnen Typus nicht. Bei
einem Pseudoreflex kann der "Reiz" die Form einer einzelnen Eigenschaft
besitzen, aber dies ist nicht möglich in den beiden wahren Typen.
Die Stellung jedes Typus in der Ökonomie des Organismus wird bemerkt.
Skinner