An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
ISSN 1492-3173
(Return to index)
By Ivan P. Pavlov(1927)
Translated by G. V. Anrep (1927)
Posted April 2000
[Classics Editor's note: Pavlov used both square and round brackets in his texts. These have been preserved but can lead to confusions as to which insertions are the author's and which are the editor's. Page numbers, reference numbers, and the occasional "sic" have been inserted in square brackets by the Classics editor. All other insertions (e.g., on p. 31) are by Pavlov. -cdg-]
LECTURE XII
Interaction of irradiation and concentration with induction.
The last three lectures were devoted to the irradiation of excitation and inhibition in the cerebral cortex and to their mutual induction. Irradiation and concentration on the one hand, and induction on the other, were dealt with separately, as if they were completely independent. In actual fact, however, it is plain that these processes are superimposed on and interact with one another. Cases of the apparently isolated existence of irradiation and concentration of either excitation or inhibition without the simultaneous presence of induction can only be regarded as exceptional, and must be interpreted either as being an expression of some definite phase of the development of these processes, or as an individual peculiarity of the nervous organization of the experimental animal. It is probable, also, that, at least in some cases, the interpretation of experiments upon irradiation and concentration was simplified artificially, since at first the existence of mutual induction was not thought of. Moreover, in the initial phases of our investigation the whole problem presented such overwhelming and chaotic complexity that many sides of the subject were intentionally ignored. We had to disregard many points, and had to obviate specially difficult problems, replacing dogs which for some reason were unsuitable or presented complex relations difficult to trace. At the present time, however, as a result of many years' experience, special attention can be directed to any new fact or any new peculiarity observed in any individual animal, and such peculiarities now raise fresh problems for investigation.
The mutual relations of irradiation and concentration of the two nervous processes with their mutual induction are exceedingly complex, and a complete knowledge of their interrelations has not yet been attained. The data available on this subject- at present do not lend themselves to systematic arrangement, but there is sufficient material to merit consideration.
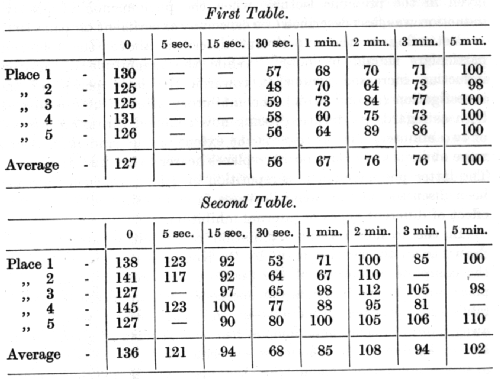
The simplest case, which will be given first, is taken from an experiment by Dr. Kreps, who worked upon the tactile analyser. [p. 205] Positive conditioned reflexes were established to tactile stimulation of two places on the thigh (1 and 2), one on the abdomen (3), one on the chest (4), and one on the shoulder (5); a place on the fore leg of the animal was completely differentiated from the others, and acted, therefore, as an inhibitory stimulus. All the positive stimuli were equalized as regards the magnitude of their effect. The actual experiments were conducted in the following manner. At the beginning of each experiment the normal magnitude of the secretory reaction for a given positive stimulus was determined ; further on in the experiment the effect of the same positive stimulus was tested again, either immediately or at various intervals of time after the termination of the inhibitory stimulus. This process was repeated with ah the rest of the positive stimuli, the whole series of experiments taking, a period of about five months for completion. The results are summarized in the two following tables, in which the magnitudes of the positive conditioned reflexes for the different places are expressed as percentages of their normal value, as determined in every experiment prior to the use of the inhibitory stimulus.
The first vertical column gives the number of the positive place of stimulation, while the upper horizontal row indicates the interval [p. 206] of time between the end of the inhibitory and the beginning of the positive stimulus. The first table represents the average figures for the total number of experiments during the whole period of investigation; the second table gives the average figures only for the last month of the investigation, the intervals of 5 and 15 seconds having been employed during the latter period only.
These experiments show that when the positive stimulus is applied immediately after the termination of the inhibitory stimulus its effect is invariably increased, when it is applied after an interval of 15 seconds its effect is below normal, and when applied towards the end of 30 seconds its effect is minimal; only towards the end of the 5th minute is complete recovery of its normal effect obtained (first table). Thus the application of the inhibitory stimulus is followed in the first place by an effect of positive induction; this is succeeded by an irradiation of inhibition, which gradually disappears so that the effect of the positive stimulus returns to normal again. A similar replacement of positive induction by an inhibitory after-effect might also have been noticed by the reader in experiments given in the previous lecture, when the phenomenon of positive induction was first described (see especially experiment on page 193).
Returning to the experiment described above, the following particulars should be noted. While the magnitude of positive induction increased somewhat towards the later period of the investigation (second table), the inhibitory after-effect diminished, both as regards its duration (being now confined within an interval of two minutes), and as regards the extent of the irradiation (being more obvious at the two positive places nearer to the inhibitory one). The latter phenomenon is a repetition of a fact which has already been discussed in the earlier lectures in connection with the after-effect of different types of internal inhibition.
In the lecture on the irradiation of inhibition attention was drawn to an observation made by Dr. Kogan, that in one of his dogs the complete extinction of the tactile conditioned stimulus in one place was always immediately succeeded by an increased excitability -- i.e. positive induction -- in the place which was furthest away from it. This phenomenon has been studied in greater detail by Dr. Podkopaev in a series of experiments in which the result of a single non-reinforcement (i.e. the first step in extinction) of a conditioned tactile alimentary reflex was investigated. Conditioned alimentary reflexes were established in a dog to tactile stimulation [p. 207] of several places on one side of the body, all of which were spaced out along a line commencing in front at the lower part of the fore limb, extending along the whole length of the trunk, and finishing at the lower part of the hind limb. All the pieces were equalized as regards the magnitude of the positive conditioned effect. Every experiment commenced by stimulating some definite place, in order to determine the normal magnitude of the reflex. The stimulation of the same or another place (at a distance of 1, 43 or 89 cms.) was now applied without reinforcement; then the particular place which had been stimulated first in the experiment was tested again at intervals of from 45 secs. to 12 mins. The experiment was concluded by testing any of the remaining places, in order to confirm the normal positive effect for the given day. Experiments were conducted with intervals of 4-5 days, in order to ensure a more or less stationary condition of the inhibition with respect to its after-effect. However, this expectation was not fulfilled, since the inhibition became more and more centred around the cortical point of its origin as the experiments proceeded, and on this account it is necessary to represent the general results in the form of three fables for three consecutive series of experiments. In the dog on which these experiments were carried out, the magnitude of the positive conditioned effect was so constant that it was possible to utilize each separate experiment instead of taking the mean of several experiments. In the tables given on page 208 the magnitude of the reflex is represented in percentages of the normal positive effect for each day. The intervals of 45 secs., 1 and 3 min. are given only in the third table, since only at this period of the experiments did the flow of saliva after administration of the non-reinforced conditioned stimulus stop in time enough to allow of a determination of the after-effect of extinction at earlier periods.
It is seen from these tables that at the beginning of the experiments (first table) the inhibition spread to the furthermost place and was still obvious 12 minutes after the extinction. Later (second table) the inhibition became limited within a distance of 43 cm., and after 4 minutes was of smaller intensity than the inhibition occurring in the preceding table after so long as 12 minutes. It can further be observed from the second table that at the remoter places there was sometimes, in the later intervals, an increase, instead of a decrease, of the effect as compared with its normal magnitude. Finally, in the last phase (third table), the inhibitory after-effect [p. 208] could be observed only in the primarily extinguished place, and this at variable periods of time, whereas in the remaining places, with the exception during the first minute of the remotest place, there was either an increase above the normal, or a return to the normal magnitude of the positive effect. The increased positive effect was most probably due to positive induction, which could be detected first at the places more remote from the starting-point of the inhibition, but only after a considerable period of time following extinction.
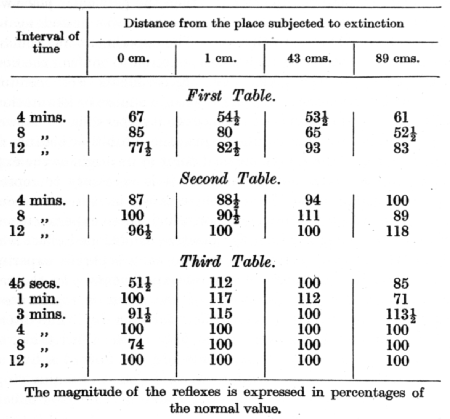
However, the effect of positive induction gradually makes itself felt at places nearer to the starting-point of the inhibition, and appears earlier after the incipient extinction. In other words, both as regards time and space, positive induction gradually overcomes and supersedes the inhibitory process.
The experiments just described present some interesting details. Firstly, they demonstrate the extreme sensitivity of the cortical elements: the effect of a single non-reinforcement exercising over a large region of the cortex a profound influence lasting for a consider able time (more than 12 minutes). Secondly, they afford a further [p. 209] example of the extreme delicacy and lability of physiological processes in the cerebral hemispheres: a relatively small influence repeated at intervals so long as 4-5 days causes a profound change in the general course of events (rapid diminution of inhibitory after-effect). Lastly, it can easily be observed that the state of different cortical points manifests a definite rhythmic undulation, both with respect to time and to distribution of inhibition. Thus in the third table the primarily extinguished place was alternately under the influence of, and free from, inhibition during 12 minutes, and a similar rhythmic oscillation was also observed in space, namely, in the distribution of excitation and inhibition in the cerebral cortex at any given moment. This fact, which is of considerable importance, will be referred to frequently in the further course of these lectures. It should be regarded as a natural result of the interaction and mutual adjustment of the two opposite nervous processes of excitation and inhibition -- just as the waves of the third degree on a blood pressure curve represent the result of the interaction of pressor and depressor influences.
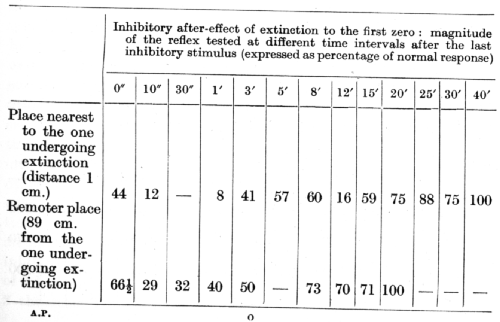
In some further experiments on the same dog, in which extinction was not restricted to a single non-reinforcement but was carried to the first zero, a, similar undulation was observed, but only at the place nearest to the one undergoing extinction. No such undulations were apparent at more remote places. The results of these experiments are presented in the following table:
 [p. 210]
[p. 210]
It is possible that the steady level of the reflex at the remoter place, which remained unaltered for 7 minutes (8th to 15th minutes), could be regarded as some expression of an undulation, especially since it corresponded to the period of a wave of inhibition at the nearest place.
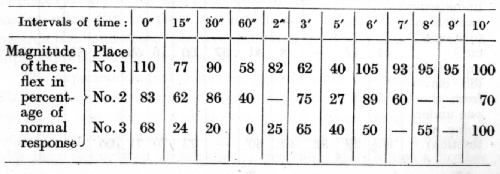
Dr. Andréev has observed similar undulations in another dog. In these experiments four apparatuses for tactile stimulation of the skin were arranged along the hind limb of the animal from the upper part of the thigh to the lower part of the leg. These places are indicated in order from above downwards by the numbers 0, 1, 2 and 3, and were spaced at equal distances of 15 cms. from one another. Stimulation of any one of the three lower places served as a positive conditioned alimentary stimulus, while stimulation of the upper place was differentiated, and acted, therefore, as an inhibitory stimulus. The differentiation, however, in this dog was not very stable. In the general course of the experiments the intervals between successive stimulations were always 7 minutes. Each experiment began with the conditioned stimulus of a buzzer or a metronome. This was followed by two positive tactile stimuli, which were applied to any of the active places, and these were followed by the positive tactile cutaneous stimulation which was being specially tested in the given experiment. All these stimuli were applied with intervals of 7 minutes. The uppermost apparatus evoking inhibition was used next. After this, and this time at different intervals varying in the different experiments from 0 to 10 minutes, the tactile stimulus which immediately preceded the inhibitory stimulus was again applied and the inhibitory after-effect determined. The following table represents the results of these experiments, the magnitude of the reflexes being calculated as a percentage of the normal positive effect, for which the mean value of the three tactile cutaneous reflexes which preceded the inhibitory stimulus was taken. Each figure given in the table is an average of three observations.
 [p. 211]
[p. 211]
The complex relations occurring in these experiments can be more readily followed on the following chart:
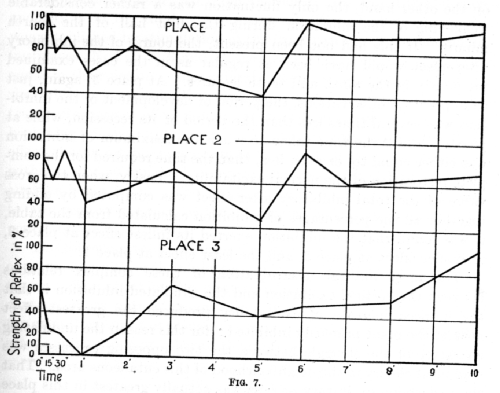
It is evident that the effects upon the different positive places of the irradiating inhibition initiated by the stimulation of place No. 0 showed more points of variance than of resemblance. The chief point which they had in common was the following: the inhibitory after-effect in all three places sooner or later reached a maximum and then disappeared. A further point in common was that all three places simultaneously revealed two waves of inhibition, the crest of one occurring at an interval of sixty seconds following the termination of the inhibitory stimulus and the second after an interval of five minutes. Here the resemblance ended; all the other features presented considerable differences. At the place nearest to the inhibitory one there was, immediately on the termination of the inhibitory stimulus, a slight increase of excitability, i.e. an effect of positive induction. In the remaining two places the inhibitory after-effect was revealed immediately. The second wave of inhibition at places 1 and 2 was greater than the first, but at place 3 the second wave was expressed only slightly, while during the [p. 212] first wave the inhibition was complete. Moreover, numerous secondary fluctuations were observed at places 1 and 2. At place 3, on the other hand, the only fluctuation was a rather considerable weakening of the inhibition during the first half of the fourth minute. If this had not been present, the course of the inhibitory after-effect would have been as regular as in the cases examined previously in the ninth and tenth lectures. At place 3, again, just as in previous observations, the period of development of the inhibition was several times less than the period of its recession, while at places 1 and 2 the time taken to reach the maximum of inhibition was either equal to, or even less, than the time required for the complete restoration of the normal excitability. Lastly, when the gross value of the fetal inhibitory after-effect; was computed by adding together all the percentages of inhibition calculated from the table, it was found that the inhibition exerted its fullest effect at place 3, a smaller effect at place 2, and the least effect at place 1.
The moat natural interpretation of these complex relations between the cortical excitation and the irradiated inhibition is that the induced increase of excitability produced the greatest effect nearest the point primarily inhibited; for this reason the undulating character of the struggle between the two opposing processes was seen most clearly in the neighbourhood of this cutaneous area. That the underlying excitatory process was actually greatest in this place is obvious from the definite result of positive induction which was observed immediately on the termination of the inhibitory stimulus at this point alone. In remoter places the inhibition was exhibited in a practically unimpeded form, first increasing gradually in strength, and then undergoing a prolonged weakening until the normal positive effect was restored. It must be noted, however, that the individual figures which were averaged exhibited for the first place variations much wider than for the intermediate place, and still wider than those for the remotest place. It is possible that the minor differences between the curves for the different places should to a certain extent be attributed to this. On the other hand, it is possible that the inconstancy in the separate figures was in itself a true expression of the greater fluctuations of the excitability in the nearest place, as compared with the remoter ones.
The following experiments by Dr. Podkopaev, which were performed on a different dog, give another striking illustration of the great complexity of the relations in question. Apparatuses for [p. 213] tactile stimulation of the skin were arranged in linear order along one side of the body of this dog as follows : No. o on the front paw, No. 1 on the shoulder, No. 2 on the chest, No. 3 on the abdomen, and No. 4 on the thigh. A positive conditioned alimentary reflex was established by stimulation of the front paw (No. 0); the remaining places (Nos. 1, 2, 3 and 4) were rendered inhibitory, and made equal in their action. Immediately after the application of any of the inhibitory stimuli, the place producing a positive reflex was tested, ifs normal effect having previously been determined at the beginning of the experiment. The experiments to determine the effect of each inhibitory place upon the positive ones were performed without any stereotyped order in the choice, first one and then another being taken. The results cannot, therefore, be attributed to any regular changes in the experimental conditions during the course of the experiments.
The following table contains the data for each of three separate experiments with each inhibitory stimulus, together with the mean figures. The effect of the positive stimulus is expressed as a percentage of its normal magnitude. It should be remembered that the excitatory effect of place No. 0 was in all cases tested immediately after applying one or other of the inhibitory stimuli.
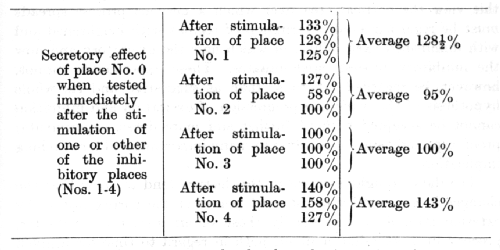
This table demonstrates clearly that the intensity of excitation at the positive place varies considerably, depending on which of the inhibitory stimuli was previously applied. When the nearest or the remotest place was the one previously stimulated, a definite effect of positive induction was readily observed: the secretory activity increased considerably as compared with the normal, the [p. 214] latent period became shorter, and frequently the alimentary motor reaction became increased in intensity. Moreover, the effect of positive induction resulting from stimulation of the remotest place was somewhat greater than that resulting from stimulation of the nearest one. No influence was observed from stimulation of place No. 3, and the effect of stimulation of place No. 2 showed no regularity, being at one test inhibitory, at another test giving rise to positive induction, and at a still further test giving neither effect. The similarity between the figures for the three separate determinations in these experiments (excluding those for place No. 2) was so great that there can be no grounds for doubting the accuracy of the average figures, and the figures relating to place No. 3 can be taken, therefore, as evidence of the existence of an intermediate zone in which neither the inhibitory after-effect nor induction was in the ascendant.
We must now consider what this experiment teaches us with regard to the intimate functional activity of the cortex. If it is assumed that the linear arrangement of the places on the akin corresponds with a similar arrangement of projection points in the cortex, the result obtained above should be regarded as striking evidence of an undulatory distribution of cortical excitability. On this view the cortical area over which a nervous process spreads must be regarded as an alternation of regions with heightened and with diminished excitability, in which now the excitatory and now the inhibitory process predominates. These experiments cannot, however, be regarded otherwise than as suggestive, and the whole hypothesis of the wave-like progression of excitation and inhibition cannot be accepted without further and more direct experimental proof, especially since it involves so many important and far-reaching implications.
The data brought forward in this lecture tend to show that the changes in excitability of different points of the cortex caused by external stimuli, and especially those of an inhibitory character, proceed in a wave-like manner, both in regard to time for a given single place, and in regard to space for different cortical points simultaneously. There is nothing surprising in this phenomenon, considering that the spread of the nervous processes on the one hand and their mutual induction on the other are established facts. Considerable difficulties arise, however, when a general rule governing the interrelations between the two processes is looked for. At [p. 215] present we are still confronted with a series of unrelated facts, which do not explain why in some cases these undulations are present, in others absent; why in some cases the spreading inhibition is preceded by a positive induction, and why in others the latter is not apparent. The available experiments, however, indicate that this variability is determined by the co-operation of three factors. These are, firstly, the functional peculiarities of the central nervous system of the experimental animal employed; secondly, the different stages of establishment of new connections in the cortex under the influence of external stimuli; and thirdly, the form in which such connections are established, since, for example, the different types of internal inhibition are known to differ considerably as regards their intensity and stability. One of the most important problems in the future complete analysis of the relations in question must be the exact determination of the part played by each of these factors in the cortical activity at any given moment, taking into consideration the relative intensities of the nervous processes involved. Of course, the problem can to some extent be dealt with even at present. In the experiments which were described a tentative approach to a solution of this question could be noticed. This is even more clearly revealed in experiments which have been recently conducted and are still in progress. In the last four lectures it could be seen that the inhibition produced by a stimulus applied at a certain point spreads over the analyser, either immediately on the termination of the inhibitory stimulus or after a short preliminary period of positive induction. For the purpose of determining what happens during the actual application of the inhibitory stimulus the following special series of experiments was carried out. In these experiments other points of the same or of other analysers were tested, not on the termination of the inhibitory stimulation, as was hitherto the practice, but while the inhibitory stimulation was still in progress, i.e. the positive stimulus to be tested was, so to speak, superimposed on the background of the inhibitory stimulus. The investigation was carried out on four different dogs by Dr. Podkopaev.
The experiments with extinctive inhibition will be considered first. The dog employed was the same which served for the experiments upon the irradiation of inhibition after a single non-reinforcement of the conditioned stimulus and after extinction to zero. In the present experiments extinction of the conditioned reflex to tactile stimulation of a definite place of the skin was carried to zero. [p. 216] The degree of inhibition was tested at two places, one at a distance of 1 cm. and the other at a distance of 89 cm. The positive stimulus was applied at each place at the 30th second of action of the inhibitory stimulus and the combined stimulus was now continued during a further 30 seconds. The stimulation of the nearer place when applied upon the background of the inhibitory stimulus gave 841/2% of the secretory effect, while that of the distant place gave 88%. In other words, the effect was practically the same at both places. It will be remembered that when the same places were stimulated immediately after the termination of the inhibitory stimulus, the nearer point gave 44% and the remoter point 661/2% (page 209). In order to appreciate these results more fully, it must be noted that in the case of the simultaneous stimulation of the inhibitory, and either of the positive places, the latent period of the reflex was considerably reduced, and the alimentary motor reaction was more sharply expressed than usually. Both these facts are unmistakable evidence of positive induction at all points of the analyser, except the one specially extinguished. The somewhat diminished secretory effect from the other places should undoubtedly be interpreted as the result of an algebraic summation of their effect with the effect of the extinguished place.
In a modification of these experiments another feature of special interest was revealed. The extinction to zero produced at a definite place of the skin was obtained by repeating the conditioned stimulus without reinforcement at intervals of 2 minutes, and the process of extinction was continued to the third zero. At the third zero stimulation of another place of the skin was superimposed on the stimulation of the inhibitory place. The reflex effect obtained from the combined stimulation was in the majority of cases equal to the usual effect of the positive stimulus taken singly. Thus it follows that an intensification of inhibition still further diminished the inhibitory effect upon the surrounding areas during the time the inhibitory stimulus lasted (enhanced induction).
Another point is also clearly brought out in these experiments. As was shown previously (page 209), the reflexes at the two positive places in this dog were still 50% inhibited two minutes after extinction of another place to the first zero. But when the action of one or another of the positive places was superimposed upon the action of the inhibitory stimulus at its third zero, again two minutes after the preceding zero, there was no inhibitory after-effect from the [p. 217] preceding extinction. It must be concluded, therefore, that during the repeated application of the inhibitory stimulus the inhibition again became concentrated at the point at which the inhibitory stimulus was applied. The spread of the inhibitory after-effect has thus to be regarded as proceeding not during, but entirely or mainly after, the termination of the inhibitory stimulus; while during its action the surrounding areas of the cortex are, to all evidence, subjected to positive induction.
Similar results were obtained by Dr. Podkopaev in another dog, in which positive conditioned alimentary reflexes were established to a definite tone and to a metronome beating at a rate of 120 per minute; an inhibitory reflex based upon differentiation was established to a stimulus of 60 beats of the metronome per minute. The tone, when superimposed upon the inhibitory stimulus of the metronome, gave out of nine experiments a slight trace of inhibition once, a normal effect four times, and an effect greater than normal four times.
In further experiments on the same problem two more dogs were employed. In one of these [experiments of Dr. Golovina] positive reflexes were established to a definite tone, to a sound of a whistle, and to a tactile stimulus. Three inhibitory reflexes were also established, all based on differentiation -- one to a definite rate of a metronome, another to a tactile stimulus, and the third to hashes of a lamp. When all these reflexes were well established, experiments were performed in which each one of the positive stimuli in turn was applied on the background of one of the inhibitory stimuli. All the tests gave uniform results. A positive stimulus combined in this way gave either a normal or else a somewhat augmented secretion, and the latent period was in most cases reduced.
In the second dog [experiments by Dr. Pavlova] positive conditioned reflexes were established to the sound of a whistle, to a sound of bubbling, and to a tactile stimulus. An inhibitory stimulus based on differentiation was established to a definite rate of a metronome. In these experiments each positive stimulus was applied several times with the inhibitory stimulus. The results were somewhat different from those obtained in the first dog. Although the latent period was in most eases reduced, the positive effect of the combined stimulus was almost constantly, and sometimes considerably, smaller than that obtained with the positive stimulus used singly. In the [p. 218] second dog, therefore, the concentration of the inhibition was not so perfect.
It should be said that the last two dogs possessed functionally different types of nervous system. While in the first dog the differentiations were in general very stable and remained unaffected by the combined stimulation, in the second dog the differentiation was frequently incomplete, and was invariably still further disturbed after the experiments with superimposition.
The series of experiments described in this lecture clearly demonstrate that, on the one hand, the positive induction arising as the result of an inhibitory stimulus applied to a definite place limits the spread of the inhibitory process, and that, on the other hand, the interaction between excitation and inhibition is determined either by the phase of establishing new relations in the cortex [experiments of Dr. Podkopaev] or by the type of nervous system in different animals [experiments by Drs. Golovina and Pavlova].