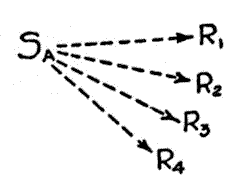
Fig. 1. The divergent excitatory mechanism. Ordinarily the different tendencies will be of widely varied strength.
Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to Classics index )
[Classics Editor's Note: Footnotes are in square brackets; references in round brackets]
First published in Psychological Review, 41, 33-54.
I. THE PRINCIPLES OF DIVERGENT AND OF CONVERGENT ASSOCIATIVE TENDENCIES
Our conception of habit is necessarily expanded with the increase in our comprehension of its phenomena. Instead of presenting a single unvarying and indistinguishable sameness, as is too often assumed, habits, even to our present meager knowledge, present a remarkably varied series of patterns. In this multiplicity it is possible, from the point of view of the present paper, to distinguish two major types. One type is characterized by a fan-like series of divergent excitatory tendencies radiating from a single stimulus, each leading to a distinct reaction. This type will be called the divergent mechanism. The other is characterized by the convergence of a number of excitatory tendencies from separate
Fig. 1. The divergent excitatory mechanism. Ordinarily the different tendencies will be of widely varied strength.
stimuli, all upon a single response. The second type will consequently be called the convergent mechanism. The divergent mechanism is represented diagrammatically in Fig. 1, [p. 34] the convergent mechanism, in Fig. 2 (13). In these diagrams and those to follow, the S's represent stimuli and the R's represent responses. Arrows with broken lines indicate acquired tendencies to action; arrows with solid lines indicate unlearned tendencies to action.
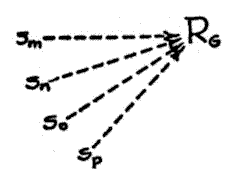
Fig. 2. The convergent excitatory mechanism.
As might readily be anticipated from the sharpness of the contrast between these two stimulus-response mechanisms, the functions performed by each in mediating adaptive behavior are characteristically distinct. Compound habit mechanisms have not been sufficiently studied from the present point of view to warrant any ultimate or comprehensive statement regarding the functional dynamics of either of the types. It is clear, however, that the first or divergent mechanism, in case the several radiating excitatory tendencies within it are antagonistic (incompatible with one another in the sense that they cannot be executed by the organism simultaneously) will give rise to a competition among themselves. This identifies the mechanism at once as the basic one responsible for the variability of response in trial-and-error learning (13). When originating in the drive stimulus (SD ), this mechanism also appears to bring about the intra-serial competition which is responsible for the serial segment elimination or 'short-circuiting' characteristic of pure-stimulus or symbolic act sequences (14). Much the same mechanism seems also to be responsible for anticipatory intrusions (22, 23), particularly of the fractional component of the anticipatory goal reaction which has been supposed to bring about the phenomenon of ideomotor action and its tremendously important functional correlate, guiding or directive ideas (15). No doubt other and equally important functions performed by this mechanism are yet to be discovered. [p. 35]
The second or convergent type of compound excitatory tendency clearly presents as a primary characteristic a response equivalence of the various stimuli involved; Sm , Sn , So , etc., are all functionally equivalent in that all alike evoke RG . The functional dynamics of this mechanism have been studied even less than have those of the divergent mechanism, but it is believed that they have far-reaching implications for the explanation of the more subtle forms of mammalian adjustment. In particular there is reason to believe that functional equivalence of stimuli plays an important role in bringing it about that habits established under certain stimulus conditions will function with little or no delay in new situations having nothing whatever as objective stimuli in common with the conditions under which the habit was originally formed.
This capacity to react appropriately to wholly new stimulus situations is of enormous biological importance because it not only saves the organism the labor of learning by trial and error to react to each new situation as it arises, but, in cases of danger where prompt and appropriate defense reactions are needed, it may make the difference between continued life and sudden death. In many such cases practice of the ordinary trial-and-error variety would be maladaptive because even one error might be fatal. This particular mechanism has an added theoretical interest because certain writers (19, p. 170). Seem to have supposed that the types of behavior mediated by it are quite impossible of explanation by any principle of association or habit action.
II. ASSOCIATIVE CONVERGENCE A MECHANISM OF AUTOMATIC HABIT TRANSFER
The power of the convergent mechanism to mediate transfer of reaction from one situation to a second which, objectively considered, may be totally different, is not difficult to show. Suppose that after the formation of the excitatory tendencies shown in Fig. 2, the combination Sm - RG has been extended by conditioning or otherwise into a sequence [p. 36] (Fig. 3) which eventuates in the terminal defense reaction Rx. Now suppose, after this has taken place, that the organism is stimulated by Sp . The sequence which would be expected to follow on the principle of ordinary habit activity is shown diagrammatically in Fig. 4. By Fig. 2, Sp , must give rise to

A chain reaction formed by extending one of the convergent excitatory tendencies shown in Fig. 2.
RG and this, in turn, to sG . But since so has been conditioned to RQ , the chain of reactions leading to RQ , will be initiated much as in Fig. 3, with the result that Sp , even though never associated with it, will bring about the terminal reaction RX . There will be the difference; however, that RQ will not have the support of the remote excitatory tendency from Sm , (14, p. 519) which it had in the original sequence (Fig. 3). [3]
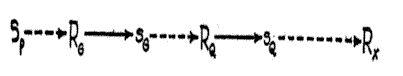
Fig. 4. Diagram illustrating the possibility of a transfer of reaction originating from one
stimulus (Sm ) to another (Sp ) which is assumed, objectively, to be totally different.
Thus we observe the superficially impossible is really quite easily conceivable without the assumption of any principles beyond the commonplace ones of conditioning or habituation. From a common-sense point of view, the explanation of the paradox lies in the fact that while, objectively considered, S m , and S p , are entirely distinct, they have, possibly by chance association, acquired identical excitatory potentialities in the body of a particular organism. This identity of excitatory capacity brings about substantially the same outcome as if they possessed an objective identity (12). [4]
The habituation transfer mechanism presented above abstractly may be clarified and emphasized by means of an experimental example. Such a case on the conditioned-reaction level seems to be furnished by a recent investigation reported by Shirley (28). Eleven human subjects were first stimulated by a faint flash of light, followed by the tap of a padded hammer against the lower eyelid. This has been shown by experiment normally to give rise to a conditioned tendency for the flash to evoke the wink. Next the subject was stimulated repeatedly by an induction shock on the fingers of one hand which, in addition to the sharp retraction of the hand from the electrodes delivering the shock, normally also gives rise to a wink. Here, then, we have two stimuli independently able to evoke the same reaction, which constitutes the convergent mechanism. This is shown diagram-

Fig. 5. Shirley's convergent conditioning situation. Here as elsewhere in the present study, the solid line
represents what is presumably an unlearned tendency, and the broken line, an acquired tendency.
matically in Fig. 5, exactly paralleling the relationship shown in Fig. 2.
After the training with the shocks, the flash was given alone and, in a small percentage of the subjects, this now evoked a finger retraction despite the fact that the flash had never previously been associated either with the shock or the finger retraction . A control experiment involving the same number of shocks similarly timed, etc., gave no finger retractions, which would seem to make it reasonably certain that the results in the case cited above were not due to mere irradiation.
The observed results of Shirley's experiment may easily be deduced from the principles of ordinary conditioning, just as was the case of transfer abstractly considered above. Since [p. 38] the shock evokes both the wink and the retraction, the proprioceptive stimuli from each should become conditioned to the other reaction either by a simultaneous, or at least by a backward (31), process, as shown in Fig. 6. Accordingly, when later the organism is stimulated by the flash, this gives
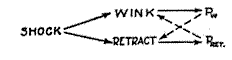
Fig. 6. Diagram showing how mutual conditioned reactions may be
set up among the several part-reactions to a single stimulus.
rise to the retraction by means of the stimulus-response sequence shown in Fig. 7. Naturally the tendency would be very weak, in part because of the specific weakness of simultaneous or backward conditioned tendencies (38), and doubt-
![]()
Fig. 7. Diagram showing the hypothetical manner in which the flash was able to evoke the
finger retraction with which it had never been associated (Shirley's 'Group C').
less also in part because the transfer involved the breaking up of a stimulus pattern.
III. ASSOCIATIVE DIVERGENCE AND CONVERGENCE THE BASIS OF THE HABIT-FAMILY HIERARCHY
With this introduction to the mechanisms of divergent and convergent excitatory tendencies, we may now turn to our main concern, which is a higher synthesis obtained by combining the two into a single functional unit. The synthesis in question is represented diagrammatically in Fig. 8. It will be observed that this figure begins with the divergent mechanism of Fig. I, and ends with the convergent mechanism of Fig. 2. Extending between the two and connecting the SA with the RG are a number of distinct behavior sequences which are supposed to differ greatly from one another both as to the qualitative nature of their activity and as to length of sequence, the first sequence (initiated by s1 ) being shortest, the second and third progressively longer, and the fourth longest of all. [p. 39]
It is to be expected that the adaptive potentialities of the mechanism obtained by combining the two dynamic tendencies discussed above should differ in certain respects from those manifested by either alone. Whereas the one mechanism by itself mediates alternative reactions, and the other brings about transfer through equivalence of reaction potentiality, there emerges from the combination (Fig. 8) the sig-
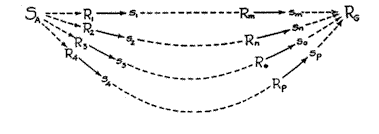
Fig. 8. The habit family hierarchy which consists of combining the divergent and the convergent excitatory mechanisms shown in Figs. 1 and 2. The length of the sequences represents inversely the excitatory strength of each at point S A . It is probable that this mechanism is related to Tolman's 'means-end-held' (33, p. 177)
nificant novel phenomenon of the habit family . A family of habits may be defined in general as a group of two or more habit sequences, all of which may be initiated by a particular stimulus and terminated by a particular reaction. [5] Despite the 'emergent' nature of the habit-family mechanism, it nevertheless retains clear functional evidences of its origin. The individual habit sequences of a particular family are still alternative in that but a single member can be active at once. At the same time they are equivalent in that all are alike in bringing about substantially the same final reaction or adjustment to the problem situation presented by SA .
Perhaps the simplest and most obvious examples of habit families are found in alternative locomotor sequences through space. If, while following a footpath we encounter a pool of water, the problem may be solved by detouring in either the right-hand or the left-hand direction. The respective behavior sequences are alternative in that we cannot go in both [p. 40] directions at the same time. They are equivalent in that each without distinction brings us again upon our path.
The alternative-pathway maze, represented conventionally in Fig. 9, gives rise to what, from the point of view of simplicity, is a limiting case of the habit-family hierarchy. As in the diagram, it is usual for such mazes to have one path somewhat shorter than the other. On the basis of trustworthy experiment (2), it is known that the shorter of the two paths, presumably in conformity with the principle of the goal
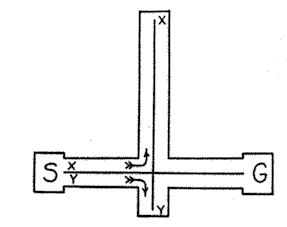
Fig. 9. Example of a simple habit family hierarchy which has been investigated experimentally. Path Y is the preferred member of the hierarchy.
gradient (16), will come to be preferred by animals which have become familiar with both. The fact that both of the action sequences begin with the same stimulus (S) and terminate in the same reaction (G) constitutes them a habit family; the fact that one sequence is preferred above the other constitutes them a hierarchy. By increasing the number of pathways and keeping the pathways all of a different length, there obviously may be generated in an animal fairly large habit-family hierarchies.
It should be especially noted, however, that the principle of habit-family hierarchy is not limited to the field of spatial orientation here emphasized. It is operative in all situations wherever there is more than one distinct action sequence which will lead to the attainment of a particular goal or subgoal. It is believed, for example, that the habit-family hierarchy constitutes the dominant physical mechanism which mediates [p. 41] such tests of truth and error as organisms employ -- that it provides the basis for a purely physical theory of knowledge. [6]
IV. THE HABIT-FAMILY HIERARCHY AND THE AUTOMATIC TRANSFER OF PRACTICE EFFECTS
The deductive explanations of maze phenomena which will be attempted in the following pages rest, for the most part, upon the following two related hypotheses:
1. That in the albino rat, habit-family hierarchies based primarily on locomotion are set up very early in life, presumably as the result of locomotion in free space, with the result that the animal at the beginning of a maze experiment is already in possession of a vast repertoire of equivalent but fairly distinct locomotor habits, any one of which, in free space, would mediate a transition of his body from the starting point to the goal .
2. That when one member of a habit-family hierarchy has attained a goal in an objectively novel situation, the learning thus acquired is transferred without specific practice to the remaining members of the hierarchy.
A crucial experimental test of the incidental assumption in the first hypothesis (that locomotor habit-family hierarchies are acquired by early running in free space) is difficult. Some general indications as to its validity might be obtained, however, by determining whether very young rats, e.g. , animals under ten or (possibly) twenty days of age, would manifest less strongly the phenomena to be expected on the assumption of the truth of the hypothesis than would older animals. Another test, though again not a crucial one, might be made by determining whether rats raised in total darkness, suspended in a hammock through which their legs would protrude so as to touch a miniature treadmill, say(thus allowing free movement and a certain amount of exercise but no genuine transit in space), would show as strong evidences of the existence of a habit-family hierarchy as would animals of the same age reared under ordinary conditions. The hypothesis leads to the expectation that they would not. By similar reasoning, animals reared in [p. 42] very small cages would show less evidences of orientation than animals raised in a cage as large as an ordinary room.
The validity of the second hypothesis must be judged in the main by the extent to which the deductions based primarily upon it agree with the facts of mammalian behavior as the latter shall finally come to be known. It is exactly this type of evidence which supports such belief as we have in molecules, atoms, and electrons.
It may be stated at once, however, that this hypothesis is definitely not put forward as an ultimate principle. On the contrary, it is confidently expected that in case it turns out to be sound, it will be found to be deducible from certain still more general principles. [7] As an illustration of the general course which this deduction (or perhaps more properly, reduction ) is likely to take, the following sketch is offered. The principles are fairly simple and the most of them are reasonably well authenticated.
From the nature of the habit family, it follows that in it all behavior sequences terminate at the same point, or in the same final act. This will often be consummatory in nature: Now there is considerable evidence that terminal reactions have a strong tendency to come forward in their entirety, as shown by abortive intrusions into behavioral sequences, which sequences were originally antecedent. Not only this, but there is evidence (29; 30) that reactions some distance from the goal also tend to come forward in the behavioral sequence, though these latter reactions appear to possess less strength in this respect than do goal reactions or reactions closely antecedent to the goal reactions. There is reason to believe, however, that in case the goal reactions or pre-goal reactions as a whole are in physiological conflict with the sequence properly leading to them, some portion of the goal reaction which chances not to be in such conflict will be temporarily detached from the goal aggregate and will move forward in the sequence independently and rather freely (15).
As yet this subject has been little investigated experimentally, so that the extent and variety of such fractional [p. 43] anticipatory reactions is very imperfectly known. Fortunately, salivation, a well-known type of anticipatory reaction, has been extensively studied. It is of significance for the understanding of the probable mechanism mediating transfer in the habit-family hierarchy that the amount and quality of anticipatory salivation is known to vary widely according to the adjustment situation (25, p. 184). Much less is known experimentally about fractional anticipatory sex behavior, though clearly this species of behavior manifests not only complex secretional activities but extensive and profound circulatory changes as well. Tumescence may be mentioned as a characteristic fractional anticipatory sex reaction. In addition to these rather gross and obvious forms of fractional anticipatory behavior, it is probable that rich variety exists, of which we as yet know little or nothing. It is conceivable, for example, that the mere visual stimulation by a light pattern may evoke hidden but characteristic reactions in the organism which could play the anticipatory role. At all events, it is not purely gratuitous to assume that both goal reactions and pre-goal reactions may possess characteristic fractional components which will be drawn forward to the beginning of all action sequences originally leading to them, and which, after each family has attained a moderate degree of integration, will be present everywhere throughout all members of a given family. [8]
It thus seems probable that the fractional anticipatory goal reaction (rG , Fig. 10) is the major mechanism which brings about the integration of the habit-family hierarchy. It is this mechanism which appears to be mainly responsible for the important and characteristic phenomenon of the transfer of practice effects from a less preferred member of a habit hierarchy to a more favored one. There is also reason to believe that under certain circumstances the fractional anticipatory goal reaction will be powerfully supported in this action by the drive stimulus (SD , Figs. 10 and 11) in, [p. 44]
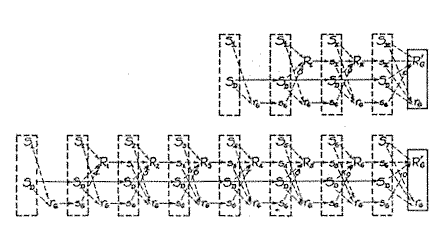
Fig. 10. Diagramatic represention of the more important excitatory tendencies involved in two behavior sequences belonging to a habit-family heirarchy. Numbers corresponding to a conventional rectilinear goal gradient are inserted on the various lines representing excitatory tendencies from s G to the several R' s ; It is noted that s G leads to R I with a strenth of 8 units and to R 1 with a strength of only 4 units. The repetition of S 1 at the outset indicates a hesitation of the animal until S 1 has had time to arouse r G .
[p. 45] case the latter is the same in the new situation as in the old. Even in cases where the drive may be superficially distinct, there is some reason to believe that there may be an innercore of identity which, if present, would doubtless aid considerably in mediating a transfer. While the inadequacy of our knowledge about these matters is such that we must be very tentative at present as to the precise details of the processes involved, it may be worth while to give an account of how the vicarious transfer under consideration might conceivably take place.
Let us suppose that the reaction sequences represented in Fig. 10 correspond to the first and last members respectively of the habit-family hierarchy shown in Fig. 8, as in their original setting. The S's represent external stimuli; the R's , the goal reactions; the rG's represent fractional anticipatory goal reactions; So, the persisting drive stimulus, e.g. , hunger; and the s's represent proprioceptive stimuli from preceding reactions. The final or goal reaction is represented as consisting of two components, R'G and rG . The latter is assumed usually to be relatively insignificant in absolute magnitude but, because of its supposed ability to move forward rather freely in the behavior sequence, to be of the greatest significance as the physical substance of goal or directive ideas. Solid arrows indicate what are presumed to be unlearned sequences, and broken lines terminating in arrows indicate tendencies to action which have been acquired by association or conditioning.
Suppose that the organism possessing the combination of excitatory tendencies shown in Fig. 10 should find itself in a somewhat novel situation presenting an initial external stimulus S1', and a different drive (SD') as well. [9] Suppose, further, that by trial-and-success the organism finds its way to the goal over the route involving the action sequence [p. 46] R1, R2, R3, R4, R5, R6, R7, RG. Now, if this action sequence is repeated a few times, rG will become conditioned to SD' and so will be brought forward to the beginning of the series (15). By virtue of this fact, rG will be brought into the presence of S1 and will therefore become conditioned to it (Fig. 11). Accordingly, whenever the animal finds himself in the presence of S1', particularly if there is a pause before the beginning of locomotion, S1' as a stimulus will evoke rG , which will bring with it the proprioceptive stimulus sG . But sG , by virtue of previous
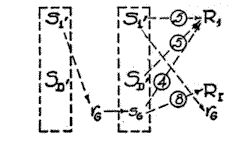
Fig. 11. Diagrammatic representation of the competition between the excitatory tendencies leading to R 1 and R I upon the transfer of the habit system shown in Fig. 10 to a new situation permitting the same behavior adjustments but having different external stimuli and also a different drive (SD'). S1' indicates the initial external stimulus complex characteristic of the new situation. It is also assumed that the goal was attained the first few time exclusively by the longer path, i.e., the one beginning with act R1 , and that during these repetitions incipient excitatory tendencies were setup leading to R1 from S1 ' and SD ' .
conditioning (Fig. 10), possesses excitatory tendencies both to R1 and to RI . Since, assuming a simple goal gradient, R1 has at this point a strength of 4 units (Fig. 10) and R I a strength of 8 units, this competition of excitatory tendencies will eventuate in reaction RI, which has never before taken place in this (objective) situation . We accordingly have here a mechanism adequate to produce the transfer of practice effects which we have been seeking. [10]
Perhaps as good an experimental example of the concrete working out of this principle as any is found in a minor detail of a study by Johnson (18, p. 37). In this portion of Johnson's experiment, two dogs, blind from the first few days of life, [p. 47] together with one normally seeing dog, were trained to open the door of a box containing food, by lifting a latch. The door of the box during the training period was always situated at the northwest corner of the box. After all three dogs had completed the learning in the original position, the box was turned 90° to the right. When the dogs had adjusted themselves to this position, the box was again turned to the right 90°;. Thus the door was opened by the dogs successively at the N.W. corner, the N.E. corner, and the S.E. corner. One of Johnson's diagrams seems to indicate that the dogs always approached the box from the south. It was observed that when the door of the box was in the third or S.E. position the two blind dogs, even after 44 and 47 successive trials, always approached it by the roundabout way of the north-This was the course by which west and the northeast corners, box had originally led them. The seeing dog, on the other hand, took the direct route to the S.E. corner after the seventh trial.
The present hypothesis would explain these observations by supposing that in the free locomotion of the seeing dog there had been developed an effective hierarchy of locomotor habits in which the shorter path possessed a distinct preference. It required seven repetitions by the non-preferred member of the hierarchy to condition the anticipatory terminal reactions to the stimuli present at the point of choice. When this had taken place, however, the sG at once evoked the dominant sequence of taking the shorter path to the S.E. corner, i.e. , practice effects acquired in a previous situation were first transferred to a "On-preferred member of a simple hierarchy by means of practice and then automatically transferred to a Preferred member of the hierarchy without any additional practice whatever in the latter act. It seems reasonable to suppose that the blind dogs, because of their sensory limitation, had not been able during their previous lives to build up such effective locomotor habit hierarchies as the seeing dog was able to do, which would account for their failure of transfer at this point in the experiment. However, these animals on still another turn of the [p. 48] box to the right, which naturally brought about a still greater difference between the two paths to the door, succeeded rather tardily in making the transfer, which indicates that they possessed the habit-hierarchy mechanism at least to a limited degree.
V. THE HABIT-FAMILY HIERARCHY AND THE PRINCIPLES OF FRUSTRATION AND DISINHIBITION
It must now be noted that the situation as represented in the preceding section will normally present certain significant complications not there. discussed. In the process of conditioning S1' to rG , additional conditioned excitatory tendencies, which may be represented by S1' -- R1 , would naturally arise. Now such excitatory tendencies must be in direct competition with R I , and so are opposed to the transfer under consideration. If, however, the conditions of the experiment are such as to leave these excitatory tendencies fairly weak, e.g., as shown in Fig. 11, the strengths of the respective excitatory tendencies might sum up as follows:
R1 = 4 + .5 + .5 = 5, RI =8
Under these conditions the excitatory tendency to RI would still be clearly dominant and the transfer would take place, though somewhat less promptly and with somewhat less certainty than without this complication.
If, on the other hand, a considerable amount of training should be given to the non-preferred member of the habit-family hierarchy under conditions such as that shown in Fig. 12, in which the preferred path is an integral part of then on-preferred path, as was the case in the experiments by Higginson, Gengerelli, and Valentine (36), a rather different situation arises. It is understood that during the main part of the training the door at E is always closed when the animals pass it on the way to D, but it is open when they return so that they can enter and go to the food at F. Under such conditions the excitatory tendencies emanating from S1' and sD' (see Fig. 11) may become fairly strong. Moreover, once [p. 49] the long path has been traversed a number of times, transfer should take place to the preferred member of the hierarchy, at which time the animals will begin attempting to take the short path to P even though the door at E is closed. But under these circumstances we should expect to have a dominant excitatory tendency undergoing frustration, i.e., being prevented from functioning. Under such conditions,
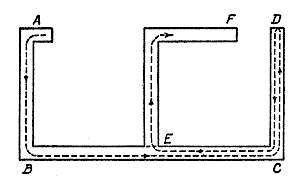
Fig. 12. Diagram of Valentine's maze (16). The broken line represents the required path of the rat during the early trials. The door at E was closed when the rat passed from B in the direction of C , but when he returned from D it was always open. F represents the point at which food was given.
there is reason to believe that an 'internal' inhibition will develop which is substantially like the experimental extinction observed in conditioned reactions. [11] We shall let the inhibitory tendency be represented by a- 1 and the two positive excitatory tendencies, one from S1' and the other from SD', each by a+1. With a moderate amount of training, then, the following situation might result:
R1 = 4 + 1+1 = 6,
RI = 8 - 1 = 7.
Even here the animal would still usually take the short path if the door were open on his way down from B . This actually took place in the experiments by Higginson (8) and Gengerelli (6).
But suppose that the training to go into the blind D (Fig. 12) were very great. Then one might expect the positive [p. 50] tendencies to R 1 to be greater and the inhibitions of the tendency to go to R I to be more profound, in which case the excitatory tendencies might sum up as follows:
R1 = 4 + 1 + 1 + .5 + .5 = 7,
RI = 8 - 4 = 4.
In such an event the animal would be expected to continue to follow the longer path unless the advantage possessed by it should be upset in some way. As a matter of fact, Valentine's animals did continue to take the longer path with fair regularity for a long time, though finally all came to take the shorter path.
But how are we to explain the fact that Valentine's animals all finally learned to take the shorter route? It might be supposed, for example, that once an animal has practiced the longer, naturally less preferred, path until the excitatory tendency to it has become dominant, he would always continue to follow it and would never thereafter eliminate the blind alley, succeeding practice serving to fixate the blind still more firmly rather than to eliminate it. This paradox, like so many that have arisen in the theory of learning, comes from assuming that only one principle, or at most a very few principles, are operating in the learning process at one time.
It happens that in this particular case, a principle from the conditioned reflex literature offers a ready explanation. It is well known (25, p. 44) that when an extra (disturbing) stimulus (or its after-effect) impinges on an excitatory tendency, the latter will be weakened; and when such a disturbing stimulus impinges on a situation where a previous excitatory tendency has suffered extinction, the inhibition becomes temporarily disinhibited, which enables at least a considerable portion of the excitatory tendency previously active to become active again (25, p. 49 ff. ). We have already presented reasons for believing that there should arise such an inhibition of the tendency to go into alley F when coming down from B .
Let us suppose that we have one of Valentine's animals which is taking consistently the long path, i.e. , that the rat [p. 51] goes from A to B , past the open door at E to C , then to D , back by C , through E and up to F . Suppose, now, that just before point B is reached (on the way to C) a loud buzzer is sounded. The excitatory tendency to C will be weakened according to the principle of external inhibition , whereas the inhibited tendency to go into F will suffer disinhibition , the latter reanimating the excitatory tendency to go directly to F The respective tendencies which were originally as appear on p. 49 would be transformed somewhat as follows: [12]
R1 = 4 + 1 + 1 + .5 + .5 = 7 - 2 = 5,
RI = 8 - 4 = 4 + 2 = 6.
The excitatory tendency of R1 which stood at 7 is weakened by two points through external inhibition, leaving 5, whereas the strength of RI, previously weakened by experimental extinction from 8 to 4, recovers two points through disinhibition so that it now stands at 6. The tendency to RI consequently becomes dominant. Therefore, after a disturbing stimulus, the animal might be expected to drop the blind into which he has been trained to go, [13] i.e., to take the turn to the left even though previously he has always entered this alley by making a turn to the right.
A second deduction which is of considerable potentiality as an experimental test of the above hypothesis depends upon the well-known fact (25, p. 65; 32) that external inhibitions and disinhibitions are notoriously transitory. This means that [p. 52] under ordinary conditions a shortly succeeding test carried out like the above but without the buzzer should be about as likely to result in the entry of alley D as were those trials made previous to the trial with the buzzer, except for the positive conditioning influence of taking on this one occasion the shorter path from A to F .
The application of these principles to Valentine's experiment is obvious. Even under the most favorable experimental conditions, slight disturbing stimuli are constantly impinging on the experimental animal; they may come from the environment, or they may originate within the animal's own body. An obvious source of external inhibition would be the open door which the animal is not accustomed to see when on his way from B , especially if a strong light should be shining through it. Other sources of disturbance might be slight sounds, odors, the bites of parasites, etc. These disturbing stimuli should produce the combination of external inhibition and disinhibition exactly as described above, and thus bring about occasional choices of the shorter path.
Since external inhibition in its various forms is inherently transitory, such short-path choices are likely to be followed by runs on the long path, particularly if the interval between trials were as much as five minutes. Thus there might be expected to take place an irregular alternation between choices of short and long paths. Since it is probable that the experimenter would not be able to sense the majority of the stimuli disturbing the animal, particularly those of internal origin, these alternations would appear to ordinary observation to take place in an entirely fortuitous manner.
REFERENCES
1. Dashiel, J. F., Direction orientation in maze running by the white rat, Comp. Psychol. Monog ., 1930, 7, No. 32.
2. De Camp, J. E., Relative distance as a factor in the white rat's selection of a path, Psychobiol. , 1934, 2, 245-253.
3. Dennis, W., A study of learning in the white rat, J. Genet. Psychol., 1934 37, 294-308.
4. Ebbinghaus, H., Memory (trans. by Ruger), New York: Teachers Coll., Columbia Univ., 1913 (original date, 1885).
5. Evans, C. L., Recent advancer in physiology, 4th ed., Philadelphia: P. Blakiston's Son and Co., 1930.
6. Gengerelli, J. A., Preliminary experiments on the causal factors in animal learning, J. Comp. Psychol., 1928, 8, 435-457.
7. Hall, M. E., Remote associative tendencies in serial learning, J. Exper. Psychol ., 1928, 11, 65-67.
8. Higginson, G. D., Visual discrimination in the white rat, J. Exper. Psychol ., 1926, 9, 337-347.
9. Hilgard, E. R., and Marquis, D. G., Acquisition, extinction, and retention of conditioned lid responses in dogs (to appear).
10. Hobhouse, L. T., Mind in evolution, London: Macmillan and Co., Ltd., 1926.
11. Holmes, S. J., The evolution of animal intelligence, New York: Henry Holt and Co., 1911.
12. Hull, C. L., Quantitative aspects of the evolution of concepts, Psychol. Monog ., 1920, 28, No 123.
13. ___, Simple trial-and-error learning: a study in psychological theory, Psychol. Rev ., 1930, 37, 241-256.
14. ___, Knowledge and purpose as habit mechanisms, Psychol. Rev ., 1930, 37, 511-525.
15. ___, Goal attraction and directing ideas conceived as habit phenomena, Psychol. Rev. , 1931, 38, 487-506.
16. ___, The goal gradient hypothesis and maze learning, Psychol. Rev ., 1932, 39, 25-43.
17. ___, Differential habituation to internal stimuli in the albino rat, J. Comp. Psychol ., 1933, 16, 255-273.
18. Johnson H. M., Audition and habit formation in the dog, Behav. Monog ., 1913-15, 2, No. 8.
19. Koffka, K., The growth of the mind (trans. by Ogden), New York: Harcourt Brace and Co., Inc., London: Kegan, Paul, Trench, Trubner and Co., Ltd., 1925.
20. Lashley, K. S., The effects of strychnine and caffeine upon the rate of learning, Psychobiol. , 1917, 1, 141-170.
21. Lepley, W. M., A theory of serial learning and forgetting based upon conditioned reflex principles, Psychol. Rev. , 1932, 39, 279-288.
22. Lumley, F. H., An investigation of the responses made in learning a multiple choice maze, Psychol. Monog ., 1931, 41, No. 189.
23. ___, Anticipation as a factor in serial and maze learning, J. Exper. Psychol ., 1932, 15, 331-342.
24. Maier, N. R. F., Reasoning in white rats, Comp. Psychol. Monog ., I929, 6, No. 29.
25. Pavlov, J. P., Conditioned reflexes, an investigation of the physiological activity of the cerebral cortex (trans. by Anrep), London: Oxford Univ. Press, 1927.
26. Peterson, J., The effect of length of blind alleys on maze learning: an experiment on twenty-four white rats, Behav. Monog ., 1916-19, 3, No. 15.
27. Robinson, E. W., and Wever, E. G., Visual distance perception in the rat, Univ. Calif. Publ. Psychol ., 1930, 4, 233-239.
28. Shipley, W. C., An apparent transfer of conditioning, J. General Psychol ., 1933, 8, 382-391.
29. Spence, K. W., The order of eliminating blinds in maze learning by the rat, J. Comp. Psychol. , 1932, 14, 9-27.
30. Spragg, S. D. S., Anticipation as a factor in maze errors, J. Comp. Psychol ., 1933, 15, 313-329.
31. Switzer, S. A., Backward conditioning of the lid reflex, J. Exper. Psychol ., 1930, 13, 76-97.
32. ___ Disinhibition of the conditioned galvanic skin response, J. General Psychol .,
1933, 9, 77-100.
33. Tolman, E. C., Purposive behavior in animals and men, New York and London:
The Century Co., 1932.
34. ____, Sign-Gestalt or conditioned reflex? Psychol. Rev ., 1933, 40, 246-255.
35. Tolman, E. C., and Hoznik, C. H.,'Insight' in rats, Univ. Calif. Publ. Psychol .,
1934, 4, 215-232.
36. Valentine, H. M., Visual perception in the white rat, J. Comp. Psychol ., 1928, 8, 369-375..
37. Wheeler, H. W., and Perkins, F. T., Principles of mental development, New York: Thomas Y. Crowell Co., 1932.
38. Wolpe, H. M., Conditioning as a function of the interval between the conditioned and the original stimulus, J. General Psychol ., 1932, 7, 80-103.
[MS. received July 26, 1933]
FOOTNOTES
[1] The substance of the major portion of the material contained in this paper has been used as a bound manuscript in certain of the writer's classes since June, 193I. During that time the author has received many valuable criticisms and suggestions from his students. Those of Dr. Kenneth W. Spence and of Mr. Neal E. Miller have been particularly valuable.
[3] In case the sequence of Fig. I were highly patterned (15, p. 500 note), e.g., by a large amount of over-training, the reaction shown in Fig. 4 would probably become exceedingly weak and might not take place at all, because the remote bond Sm -- RQ would be lacking in the stimulus combination shown in Fig. 4.
[4] As a result of this difference in the basic mechanism involved, it is to be expected that transfer on the basis of objective identity in the stimulus situation should be largely common to the members of a given species, whereas transfer based on acquired identical excitatory potentialities should be very unevenly distributed within a population. Such generality as the latter would show, would be based on the similarity of environmental circumstances. In some cases even this might produce considerable uniformity in transfer capacity.
[5] The term 'families' is here used in much the same sense that geometers use the term to designate a series of curves, such as parabolas, which originate at the same point but thereafter follow different courses, all being generated by a single formulab ut each having a different value for one of the parameters.
[6] This matter is reserved for elaboration in a subsequent paper.
[7] This statement also holds for the principle of the goal gradient (16).
[8] A somewhat detailed discussion of one mechanism by which goal and near-goal reactions may be displaced forward in a behavior sequence may be found in reference 14. A second mechanism has been proposed by Lepley (21).
[9] The deduction would be considerably easier if the original drive (SD) were assumed in the new situation, but the outcome would be less general than to assume a distinct drive. It may easily be deduced from these and related considerations that, other things being equal, transfer should be distinctly more effective where the same drive (e.g. hunger) is involved in both the old and the new situations. This deduction may easily be checked by experiment.
[10] It will be noted that the mechanism here indicated as mediating the transfer of habituation, while differing in important respects from that presumably responsible for the transfer in Shipley's conditioned reflex experiment, resembles it in that the element upon which the transfer was based is an art.
[11] See Part II for an elaboration of this point.
[12] It is not yet clear from the experimental work on conditioned reflexes whether a disinhibiting stimulus affects only the inhibitory component or whether its effect is also upon the excitatory component as well. It seems reasonable to assume that the latter is the case. The net gain in strength of reaction in such a case would be due to the well-known fact (25, p. 66) that inhibitory tendencies suffer much greater external inhibition from stimuli of moderate strength than do excitatory tendencies. Thus the net gain in the strength of RI of two points (from 4 to 6) might result from a combined reduction of the basic excitatory tendency of 8 to 7 through external inhibition and a gain in effective excitatory strength through a reduction by disinhibition of the four units of inhibition to 1. Thus the application of the results of conditioned reflex experiments to more complex situations give a unique significance to certain phases of conditioned reactions hitherto disregarded.
[13] Perhaps one of the most important biological functions performed by external inhibition (and disinhibition) is to prolong trial-and-error and prevent permanent fixation of maladaptive habit tendencies of which the long path in Valentine's experiment is an example.