An internet resource developed by
Christopher
D. Green
York University, Toronto, Ontario
(Return to Classics index)
THE TIME TAKEN UP BY CEREBRAL OPERATIONS.[1]
James McKeen Cattell (1886b)
Assistant in the Psychological Laboratory, University of Leipsic.
First published in Mind, 11, 377-392.
Part 3 of 4
III. The Perception-Time.
The object most quickly perceived through the sense of sight is a simple light. In order to investigate the time required I took two cards, one entirely black, the other having on the black a white surface. One of the cards, the observer not knowing which, was placed by the experimenter in the springs of the gravity-chronometer, and the clockwork of the chronoscope was set in motion. The observer fixated the grey spot on the screen immediately before the centre of the white surface (supposing this card to be there), and with his left hand broke an electric current and let the screen fall. The card appeared at the point fixated, and at this same instant the current controlling the chronoscope was closed. The observer either saw nothing, or at the point fixated a white surface. If the light appeared he lifted his hand as quickly as possible, if there was no light he did not let go the key, and the hands of the chronoscope ran on until the clockwork was stopped by the experimenter. Twenty-six experiments were made in a series, the white light occurring thirteen times. Determinations were only made when the light occurred, so the averages in this section are from thirteen reactions (in the corrected series from ten). It will be seen that, as the observer tries to make the reaction as quickly as possible, he may lift his hand when the light is not present. If this happens often the times measured are not correct, but too short, since we may assume that the observer lifts his hand as often when the white light is present before he has seen it, as he makes the motion when no light comes. We must however expect such a false reaction occasionally to occur, otherwise we might assume that the reaction is not made in the minimum time when the light is present. In these experiments such false reactions scarcely happened except when the observer was disturbed, or when the impressions to be distinguished were similar (E from F, for example). In the first case the average is not seriously affected, as the reactions are as apt to be unduly retarded as unduly hurried. In the second case false reactions lead us to suppose that some of the reactions on the stimulus are too short. The method I have introduced of giving [p. 379] a corrected average eliminates all premature reactions. I give in the Tables the number of false reactions made;[4] it would have been well if v. Kries and Auerbach, Merkel and others had done the same.
We can now examine the Table giving the time needed to perceive and react on a white surface.
The reaction was made with the speech-organs in quite the same manner. When the white surface was seen the observer said 'Weiss' and the hands of the chronoscope were stopped by means of the lip-key or sound-key. When no white surface was present the observer said nothing, and the hands ran on until the experimenter stopped the clockwork.
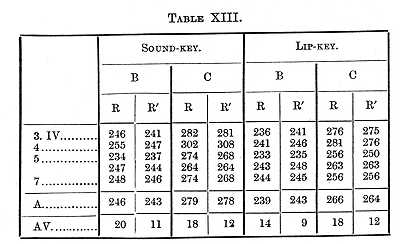
If instead of two black cards on one of which there is a white surface, we take two white cards on one of which there is a black surface, and let the observer react only when the black is present, the conditions are substantially as before; the perception may require slightly longer, the will-time is probably the same. The results of such experiments are given in Table XIV.
If, instead of black, we place a colour on the white card, the perception becomes slightly more difficult; it is not quite so easy to see that something is there when it is yellow as when it is black, the will-time however presumedly remains the same. In one series of experiments (to the left in Table XV.) only one colour was used at a time, in a second series (right in Table [p. 381] XV.) ten colours, the observer not knowing which was to come, but not needing to distinguish it before making the motion.
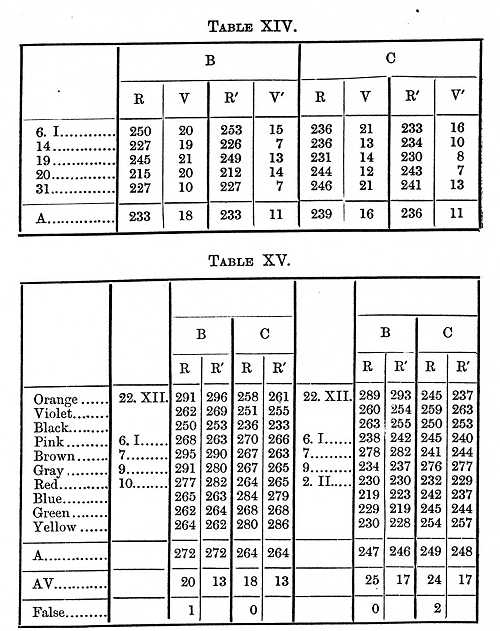
We next determine the perception-time when it is necessary to distinguish the colour. Two cases were considered; in one the colours were taken in pairs, and one colour was distinguished [p. 382] from the other; in the second each colour was distinguished from ten colours. With blue and red electric lights (the above-mentioned Puluj's tube seen through coloured glasses) I got as perception- and will-time 75s for B, 109 for C.[5] In most of my experiments however, with aid of the gravity-chronometer, I used daylight reflected from coloured surfaces, these exciting the processes with which our brain is occupied in our daily life. Red and blue and green and yellow were taken in pairs, the coloured surface being 3 x 30 mm. The numbers in Table XVI. give the average of six series.
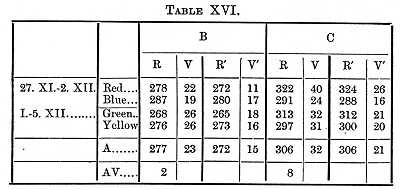
If we average together the results given in Tables XVI. and [p. 383] XVII., and subtract the reaction-time and supposed will-time, we find that it took B 100, C 110s, to distinguish one colour from another.
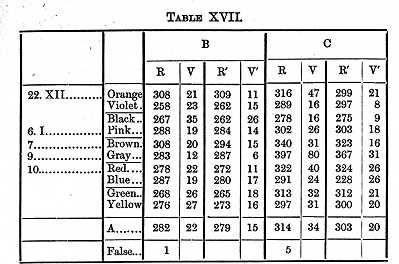
This gives as the time needed to distinguish a colour 105s for B, 117 for C; respectively 5 and 7s longer than it took to distinguish one colour from another, and 26 and 41s longer than it took to see that a colour was present when it was not necessary to distinguish it.
The results given in Table XVIII. (where the reaction was made with the hand) were obtained at the beginning of the investigation; the determinations were repeated after four months of constant practice, and again after a pause of three months, the results being given in Table XIX.
Practice therefore shortened the perception- and will-times about 30s for B and 20 for C, and this decrease in the length of the times was not lost by an interruption in the practice.
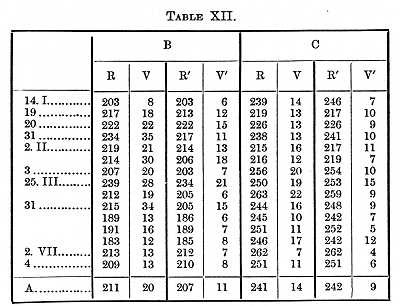
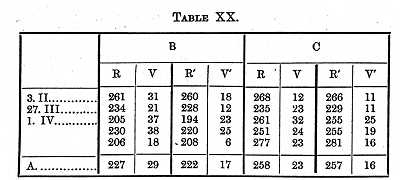
With the same methods I found the time it takes to see or distinguish a letter. I tried in my experiments to determine the time taken up by those operations which are constantly going on in the brain; the letters chosen therefore were such as we usually have to read (of the size in which this is printed). The time for larger letters is somewhat shorter. In the first experiments it was not necessary to distinguish the letter, only to know that a letter was present; the conditions were consequently the same as in the first experiments (Table XV.) on colours.
[p. 384]
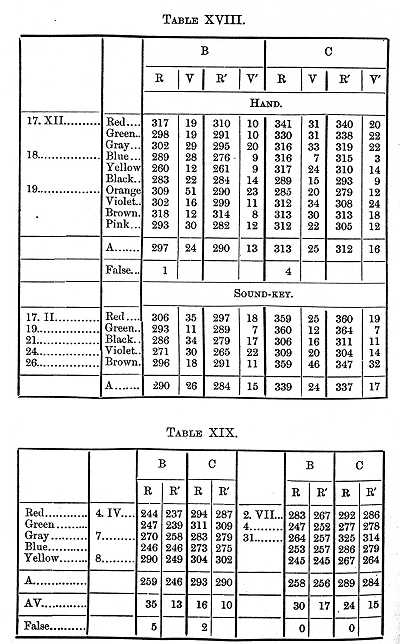
[p. 385] It therefore (making the same assumptions as above) took B 47, C 58s, to see that a small object was on a white surface.
We now come to consider the time needed to distinguish one letter from
all the others; that is the time it takes to see a letter. This is a process
with which our brain is constantly busy; the time taken up by it is therefore
of special interest. If for example the time is different for the several
letters, it is a matter of the greatest practical importance, for those
letters which it takes the longest to see might be so modified as to shorten
the time. If it takes 20s longer to see E than
it would to see a symbol that might be taken in its place, say ![]() ,
it is startling if we calculate how much time is being wasted and how much
unnecessary strain is being put on eye and brain. I have published[6]
extended series [p. 386] of experiments, determining the time the light
reflected from a printed letter must work on the retina in order that it
may be possible to see the letter. These experiments show that there is
a great difference in the legibility of the several letters; out of 270
trials W was read correctly 241, E only 63 times. In this case the whole
time was short, 1 to 1·5s, and the difference
in the time for the several letters correspondingly small. When however
we determine the entire time needed to recognise the letter, we may expect
to find the time considerably shorter for a simple and distinct symbol
than for one complicated or easily confused with others, just as the time
is shorter for a colour than for a letter.[7] The speech-organs
as well as the hand were used in these experiments. Here however a slight
complication is added, as we cannot be sure that a difference in the time
for the several letters is to be referred only to the perception-time,
it being possible that the time needed to name the several letters or to
register the different motions may be different. This difference in time
can however only be very small, as the observer knew what letter he had
to name, so there was no choice between different motions, as in the experiments
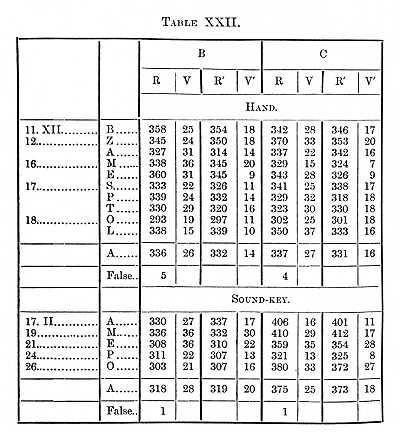
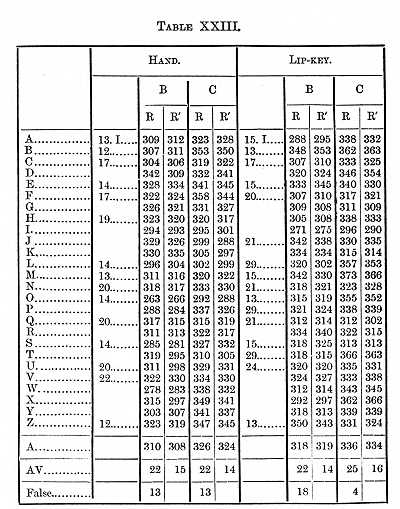
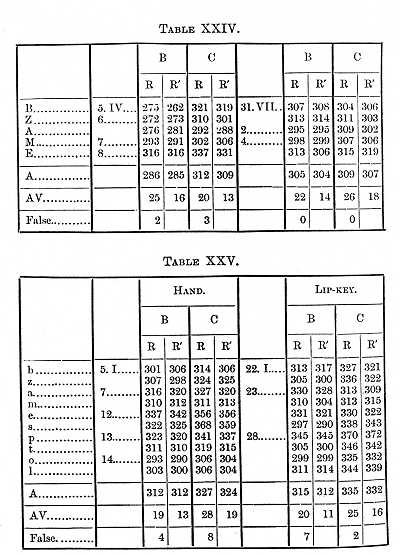
to be considered in the next section of this paper. Tables XXII.-XXIV.
placed, with others, at the end of this paper) give the results obtained
at different times, the motion being made both with the hand and the speech-organs.
,
it is startling if we calculate how much time is being wasted and how much
unnecessary strain is being put on eye and brain. I have published[6]
extended series [p. 386] of experiments, determining the time the light
reflected from a printed letter must work on the retina in order that it
may be possible to see the letter. These experiments show that there is
a great difference in the legibility of the several letters; out of 270
trials W was read correctly 241, E only 63 times. In this case the whole
time was short, 1 to 1·5s, and the difference
in the time for the several letters correspondingly small. When however
we determine the entire time needed to recognise the letter, we may expect
to find the time considerably shorter for a simple and distinct symbol
than for one complicated or easily confused with others, just as the time
is shorter for a colour than for a letter.[7] The speech-organs
as well as the hand were used in these experiments. Here however a slight
complication is added, as we cannot be sure that a difference in the time
for the several letters is to be referred only to the perception-time,
it being possible that the time needed to name the several letters or to
register the different motions may be different. This difference in time
can however only be very small, as the observer knew what letter he had
to name, so there was no choice between different motions, as in the experiments
to be considered in the next section of this paper. Tables XXII.-XXIV.
placed, with others, at the end of this paper) give the results obtained
at different times, the motion being made both with the hand and the speech-organs.
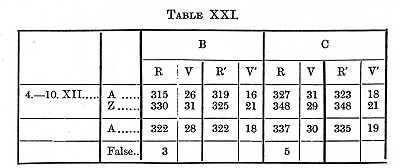
A shortening in the time through practice will be noticed in these Tables; if we take Table XXIII., which contains the most determinations and times representing about the average of the three Tables, we find the perception-time for a capital letter of the size in which this is printed to be 119s for B, 116 for C. The Tables contain the results of a great many experiments, but not enough to determine finally the time for the several letters; if however the four series made with the hand on E and M are averaged together, we find that it took B 19, C 22s longer to see E than to see M. The order for the five letters on which four series were made is M A Z B E, which (except the position of Z) agrees with the order of legibility established in the paper referred to.
Similar determinations were made with the small letters, the results being given in Table XXV. It seems from this Table [p. 387] that the perception-time is about the same for the large and small letters, which agrees with experiments I have made by an entirely different method (see MIND 41).
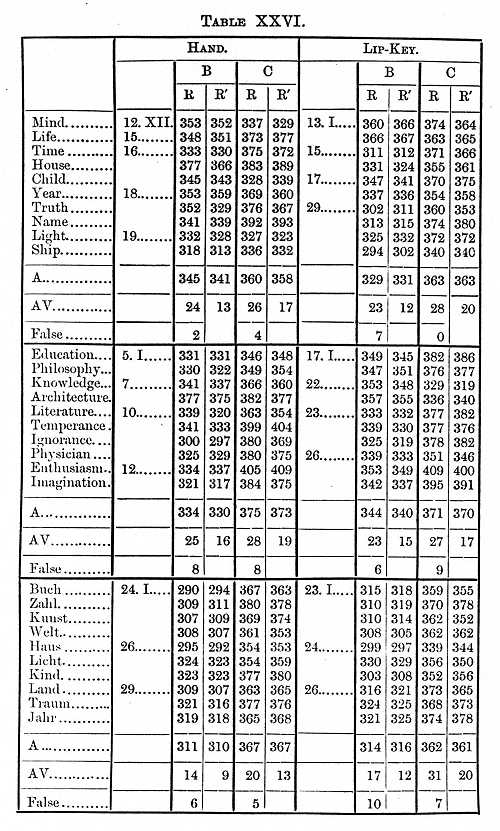
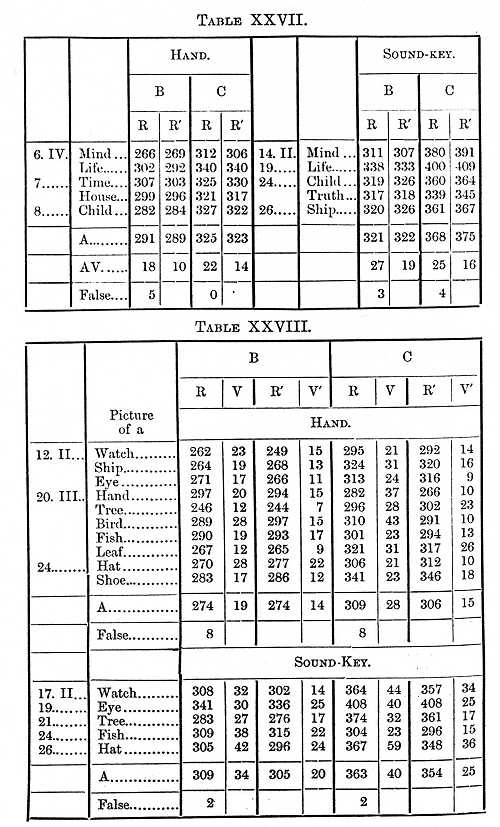
We now come to consider the time it takes to see a word, a process with which the brain is constantly occupied. Twenty-six words were taken, and when the expected one was seen the observer lifted his hand. The perception-time so determined is the time needed to distinguish the word from the other twenty-five; the time is slightly longer when it is necessary to distinguish words from others very similar in form; for example, hand from band. Indeed we must remember that perception is not a sharply defined process. As I have shown, we see a letter before we see what letter it is; in like manner a further time passes before we see the letter in all its details, that it is not perfectly printed, for example. The perception-time for a painting by Raphael is indefinitely long. The results of experiments with English and German words are given in the Tables XXVI.-VII.
The Tables give us a perception-time for short English words B 132, C 141s; for short German words B 118, C 150s; for long English words B 154, C 158s. The time was therefore slightly shorter (B 22, C 17) for a short than for a long word, and for a word in the native than in a foreign language (B 14, C 9). It will be noticed that the perception-time is only slightly longer for a word than for a single letter; we do not therefore perceive separately the letters of which a word is composed, but the word as a whole. The application of this to teaching children to read is evident; I have already in connexion with other experiments called attention to it.
The only other perception-time we have to consider is for a picture. It takes, we may suppose, about the same time to recognise the picture of a tree as it takes to see the tree itself; this is consequently a process nearly always going on in the brain. I had carefully drawn twenty-six pictures of common objects, tree, hand, ship, etc, about one square cm. in size, the method of determining the perception-time being as before.
We thus find that the perception-time for a picture, and we may assume for the objects we are continually seeing in our daily life, was 96s for B, 117 for C, about the same as for a colour and shorter than for a letter or word.
(To be concluded.)
Footnotes
[2] Physiol. Psych., ii., 247 ff.; Phil. Studien., i., 25 ff.
[3] De Jaager, De physiologische Tijd Bij psychische Processen, Utrecht, 1865; Donders, Archiv f. Anat. u. Physiol., 1868.
[4] After "false," the entire number made during the series given in the column under which it stands.
[5] These are the only experiments described in this
section which had been previously made; Donders (Archiv f. Anat.
u.
Physiol., 1868) found the time to be 184s,
Wundt (Physiol. Psych., 11, 251) 210 to 250s,
v. Kries and Auerbach, working under the direction of Helmholtz (Archiv
f. Anat. u. Physiol, 1877), 12 and 34s.
I cannot accept the results reached by these
latter experimenters. The times seem to be too short to be correct.
I do not know where the error lies, the experiments having apparently been
made with great care, but the simple reactions are very long, the reactions
with perception and volition very short. The latter may have been made
unduly short through the frequent occurrence of premature reactions (the
number of false reactions is not given); at all events I consider their
method of calculating the averages dangerous, they ignoring what reactions
they saw fit. They do not give the number of measurements made in the
series, but in the model series given in the appendix, we find that in
one 22 reactions were used, in one on the perception of light only 9; we
may therefore assume that in the latter series over half of the reactions
were ignored. If the mean variation of the reactions used in this series
be calculated, it will be found to be 6 (smaller, I imagine, than the mean
error of the recording apparatus); the mean variation of the corresponding
series of simple reactions (from which determinations had also been omitted)
is 12s. When averages are made up in this way
any results desired can be obtained.
[6] Phil. Studien, ii. 4; Brain, No. 31.
[7] I have not been able to determine accurately and
finally the perception-time for different alphabets and for the several
letters. In these experiments the different letters cannot well be used
in the same series, and further in half the cases no measurement is made.
As the difference in the times is small and the variation of the series
not inconsiderable, a large number of experiments must be made before the
difference in the time for the several letters can be determined with certainty.
This is however not only a subject of scientific interest, but also of
great practical importance; it is to be hoped that it will be thoroughly
investigated by independent experimenters.
TABLES XXII. - XXVIII.