Classics in the History of Psychology
An internet resource developed by
Christopher D. Green
York University, Toronto, Ontario
(Return to index)
The Principles of Psychology
William James (1890)
CHAPTER II
The Functions of the Brain
If I begin chopping the foot of a tree, its branches are
unmoved by my act, and its leaves murmur as peacefully as ever in the
wind. If, on the contrary, I do violence to the foot of a fellow-man, the
rest of his body instantly responds to the aggression by movements of alarm
or defence. The reason of this difference is that the man has a nervous
system whilst the tree has none; and the function of the nervous system
is to bring each part into harmonious co-operation with every other. The
afferent nerves, when excited by some physical irritant, be this as gross
in its mode of operation as a chopping axe or as subtle as the waves of
light, conveys the excitement to the nervous centres. The commotion set
up in the centres does not stop there, but discharges itself, if at all
strong, through the efferent nerves into muscles and glands, exciting movements
of the limbs and viscera, or acts of secretion, which vary with the animal,
and with the irritant applied. These acts of response have usually the
common character of being of service. They ward off the noxious stimulus
and support the beneficial one; whilst if, in itself indifferent, the stimulus
be a sign of some distant circumstance of practical importance, the animal's
acts are addressed to this circumstance so as to avoid its perils or secure
its benefits, as the case may be. To take a common example, if I hear the
conductor calling ' All aboard!' as I enter the depot, my heart first stops,
then palpitates, and my legs respond to the air-waves falling on my tympanum
by quickening their movements. If I stumble as I run, the sensation of
falling provokes a movement of the hands towards the direction of the fall,
the effect of which is to shield the body from too sudden a shock. If a
cinder enter my eye, its lids close forcibly and a copious flow of tears
tends to wash it out.
[p.13] These three responses to a sensational stimulus differ, however,
in many respects. The closure of the eye and the lachrymation are quite
involuntary, and so is the disturbance of the heart. Such involuntary responses
we know as 'reflex' acts. The motion of the arms to break the shock of
falling may also be called reflex, since it occurs too quickly to be deliberately
intended. Whether it be instinctive or whether it result from the pedestrian
education of childhood may be doubtful; it is, at any rate, less automatic
than the previous acts, for a man might by conscious effort learn to perform
it more skilfully, or even to suppress it altogether. Actions of this kind,
into which instinct and volition enter upon equal terms, have been called
'semi-reflex.' The act of running towards the train, on the other hand,
has no instinctive element about it. It is purely the result of education,
and is preceded by a consciousness of the purpose to be attained and a
distinct mandate of the will. It is a 'voluntary act.' Thus the animal's
reflex and voluntary performances shade into each other gradually, being
connected by acts which may often occur automatically, but may also be
modified by conscious intelligence.
An outside observer, unable to perceive the accompanying consciousness,
might be wholly at a loss to discriminate between the automatic acts and
those which volition escorted. But if the criterion of mind's existence
be the choice of the proper means for the attainment of a supposed end,
all the acts seem to be inspired by intelligence, for appropriateness
characterizes them all alike. This fact, now, has led to two quite opposite
theories about the relation to consciousness of the nervous functions.
Some authors, finding that the higher voluntary ones seem to require the
guidance of feeling, conclude that over the lowest reflexes some such feeling
also presides, though it may be a feeling of which we remain unconscious.
Others, finding that reflex and semi-automatic acts may, notwithstanding
their appropriateness, take place with an unconsciousness apparently complete,
fly to the opposite extreme and maintain that the appropriateness even
of voluntary actions owes nothing to the fact that consciousness attends
them. They are, according to these writers, results of physiological mechanism
pure [p.14] and simple. In a near chapter we shall return to this controversy
again. Let us now look a little more closely at the brain and at the ways
in which its states may be supposed to condition those of the mind.
THE FROG'S NERVE-CENTRES.
Both the minute anatomy and the detailed physiology of the brain are achievements
of the present generation, or rather we may say (beginning with Meynert)
of the past twenty years. Many points are still obscure and subject to
controversy; but a general way of conceiving the organ has been reached
on all hands which in its main feature seems not unlikely to stand, and
which even gives a most plausible scheme of the way in which cerebral and
mental operations go hand in hand.
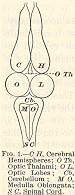 The best way to enter
the subject will be to take a lower creature, like a frog, and study by
the vivisectional method the functions of his different nerve-centres.
The frog's nerve-centres are figured in the accompanying diagram, which
needs no further explanation. I will first proceed to state what happens
when various amounts of the anterior parts are removed, in different frogs,
in the way in which an ordinary student removes them; that is, with no
extreme precautions as to the purity of the operation. We shall in this
way reach a very simple conception of the functions of the various centres,
involving the strongest possible contrast between the cerebral hemispheres
and the lower lobes. This sharp conception will have didactic advantages,
for it is often very instructive to start with too simple a formula and
correct it later on. Our first formula, as we shall later see, will have
to be softened down somewhat by the results of more careful experimentation
both on frogs and birds, and by those of the most recent observations on
dogs, [p.15] monkeys, and man. But it will put us, from the outset, in
clear possession of some fundamental notions and distinctions which we
could otherwise not gain so well, and none of which the later more completed
view will overturn.
The best way to enter
the subject will be to take a lower creature, like a frog, and study by
the vivisectional method the functions of his different nerve-centres.
The frog's nerve-centres are figured in the accompanying diagram, which
needs no further explanation. I will first proceed to state what happens
when various amounts of the anterior parts are removed, in different frogs,
in the way in which an ordinary student removes them; that is, with no
extreme precautions as to the purity of the operation. We shall in this
way reach a very simple conception of the functions of the various centres,
involving the strongest possible contrast between the cerebral hemispheres
and the lower lobes. This sharp conception will have didactic advantages,
for it is often very instructive to start with too simple a formula and
correct it later on. Our first formula, as we shall later see, will have
to be softened down somewhat by the results of more careful experimentation
both on frogs and birds, and by those of the most recent observations on
dogs, [p.15] monkeys, and man. But it will put us, from the outset, in
clear possession of some fundamental notions and distinctions which we
could otherwise not gain so well, and none of which the later more completed
view will overturn.
If, then, we reduce the frog's nervous system to the spinal cord alone,
by making a section behind the base of the skull, between the spinal cord
and the medulla oblongata, thereby cutting off the brain from all connection
with the rest of the body, the frog will still continue to live, but with
a very peculiarly modified activity. It ceases to breathe or swallow; it
lies flat on its belly, and does not, like a normal frog, sit up on its
fore paws, though its hind legs are kept, as usual, folded against its
body and immediately resume this position if drawn out. If thrown on its
back, it lies there quietly, without turning over like a normal frog. Locomotion
and voice seem entirely abolished. If we suspend it by the nose, and irritate
different portions of its skin by acid, it performs a set of remarkable
'defensive' movements calculated to wipe away the irritant. Thus, if the
breast be touched, both fore paws will rub it vigorously; if we touch the
outer side of the elbow, the hind foot of the same side will rise directly
to the spot and wipe it. The back of the foot will rub the knee if that
be attacked, whilst if the foot be cut away, the stump will make ineffectual
movements, and then, in many frogs, a pause will come, as if for deliberation,
succeeded by a rapid passage of the opposite unmutilated foot to the acidulated
spot.
The most striking character of all these movements, after their teleological
appropriateness, is their precision. They vary, in sensitive frogs and
with a proper amount of irritation, so little as almost to resemble in
their machine-like regularity the performances of a jumping-jack, whose
legs must twitch whenever you pull the string. The spinal cord of the frog
thus contains arrangements of cells and fibres fitted to convert skin irritations
into movements of defence. We may call it the centre for defensive movements
in this animal. We may indeed go farther than this, and by cutting the
spinal cord in various places find that its separate segments are independent
mechanisms, for appropriate activities of the head and of the arms and
legs respec-[p.16] tively. The segment governing the arms is especially
active, in male frogs, in the breeding season; and these members alone
with the breast and back appertaining to them, everything else being cut
away, will then actively grasp a finger placed between them and remain
hanging to it for a considerable time.
The spinal cord in other animals has analogous powers. Even in man it
makes movements of defence. Paraplegics draw up their legs when tickled;
and Robin, on tickling the breast of a criminal an hour after decapitation,
saw the arm and hand move towards the spot. Of the lower functions of the
mammalian cord, studied so ably by Goltz and others, this is not the place
to speak.
If, in a second animal, the cut be made just behind the optic lobes
so that the cerebellum and medulla oblongata remain attached to the cord,
then swallowing, breathing, crawling, and a rather enfeebled
jumping and swimming are added to the movements
previously observed.[1] There are other reflexes too.
The animal, thrown on his back, immediately turns over to his belly. Placed
in a shallow bowl, which is floated on water and made to rotate, he responds
to the rotation by first turning his head and then waltzing around with
his entire body, in the opposite direction to the whirling of the bowl.
If his support be tilted so that his head points downwards, he points it
up; he points it down if it be pointed upwards, to the right if it be pointed
to the left, etc. But his reactions do not go farther than these movements
of the head.; He will not, like frogs whose thalami are preserved, climb
up a board if the latter be tilted, but will slide off it to the ground.
If the cut be made on another frog between the thalami and the optic
lobes, the locomotion both on land and water becomes quite normal, and,
in addition to the reflexes already shown by the lower centres, he croaks
regularly whenever he is pinched under the arms. He compensates rotations,
etc., by movements of the head, and turns over from his back; but still
drops off his tilted [p.17] board. As his optic nerves are destroyed by
the usual operation, it is impossible to say whether he will avoid obstacles
placed in his path.
When, finally, a frog's cerebral hemispheres alone are cut off by a
section between them and the thalami which preserves the latter, an unpractised
observer would not at first suspect anything abnormal about the animal.
Not only is he capable, on proper instigation, of all the acts already
described, but he guides himself by sight, so that if an obstacle be set
up between him and the light, and he be forced to move forward, he either
jumps over it or swerves to one side. He manifests sexual passion at the
proper season, and, unlike an altogether brainless frog, which embraces
anything placed between his arms, postpones this reflex act until a female
of his own species is provided. Thus far, as aforesaid, a person unfamiliar
with frogs might not suspect a mutilation; but even such a person would
soon remark the almost entire absence of spontaneous motion-that is, motion
unprovoked by any present incitation of sense. The continued movements
of swimming, performed by the creature in the water, seem to be the fatal
result of the contact of that fluid with its skin. They cease when a stick,
for example, touches his hands. This is a sensible irritant towards which
the feet are automatically drawn by reflex action, and on which the animal
remains sitting. He manifests no hunger, and will suffer a fly to crawl
over his nose unsnapped at. Fear, too, seems to have deserted him. In a
word, he is an extremely complex machine whose actions, so far as they
go, tend to self-preservation ; but still a machine, in this sense-that
it seems to contain no incalculable element. By applying the right sensory
stimulus to him we are almost as certain of getting a fixed response as
an organist is of hearing a certain tone when he pulls out a certain stop.
But now if to the lower centres we add the cerebral hemispheres, or
if, in other words, we make an intact animal the subject of our observations,
all this is changed. In addition to the previous responses to present incitements
of sense, our frog now goes through long and complex acts of locomotion
spontaneously, or as if moved by what in our-[p.18] selves we should
call an idea. His reactions to outward stimuli vary their form, too. Instead
of making simple defensive movements with his hind legs like a headless
frog if touched, or of giving one or two leaps and then sitting still like
a hemisphereless one, he makes persistent and varied efforts at escape,
as if, not the mere contact of the physiologist's hand, but the notion
of danger suggested by it were now his spur. Led by the feeling of hunger,
too, he goes in search of insects, fish, or smaller frogs, and varies his
procedure with each species of victim. The physiologist cannot by manipulating
him elicit croaking, crawling up a board, swimming or stopping, at will.
His conduct has become incalculable. We can no longer foretell it exactly.
Effort to escape is his dominant reaction, but he may do anything
else, even swell up and become perfectly passive in our hands.
Such are the phenomena commonly observed, and such the impressions
which one naturally receives. Certain general conclusions follow irresistibly.
First of all the following:
The acts of all the centres involve the use of the same muscles.
When a headless frog's hind leg wipes the acid, he calls into play all
the leg-muscles which a frog with his full medulla oblongata and cerebellum
uses when he turns from his back to his belly. Their contractions are,
however, combined differently in the two cases, so that the results
vary widely. We must consequently conclude that specific arrangements of
cells and fibres exist in the cord for wiping, in the medulla for turning
over, etc. Similarly they exist in the thalami for jumping over seen obstacles
and for balancing the moved body; in the optic lobes for creeping backwards,
or what not. But in the hemispheres, since the presence of these organs
brings no new elementary form of movement with it, but only determines
differently the occasions on which the movements shall occur,
making the usual stimuli less fatal and machine-like; we need suppose no
such machinery directly co-ordinative of muscular contractions to
exist. We may rather assume, when the mandate for a wiping-movement is
sent forth by [p.19] the hemispheres, that a current goes straight to the
wiping-arrangement in the spinal cord, exciting this arrangement as a whole.
Similarly, if an intact frog wishes to jump over a stone which he sees,
all he need do is to excite from the hemispheres the jumping-centre in
the thalami or wherever it may be, and the latter will provide for the
details of the execution. It is like a general ordering a colonel to make
a certain movement, but not telling him how it shall be done.[2]
The same muscle, then, repeatedly represented at different heights;
and at each it enters into a different combination with other muscles to
co-operate in some special form of concerted movement. At each height
the movement is discharged by some particular form of sensorial
stimulus. Thus in the cord, the skin alone occasions movements; in
the upper part of the optic lobes, the eyes are added; in the thalami,
the semi-circular canals would seem to play a part; whilst the stimuli
which discharge the hemispheres would seem not so much to be elementary
sorts of sensation, as groups of sensations forming determinate objects
or things. Prey is not pursued nor are enemies shunned
by ordinary hemisphereless frogs. Those reactions upon complex circumstances
which we call instinctive rather than reflex, are already in this animal
dependent on the brain's highest lobes, and still more is this the case
with animals higher in the zoological scale.
The results are just the same if, instead of a frog, we take a pigeon,
and cut out his hemispheres as they are ordinarily cut out for a lecture-room
demonstration. There is not a movement natural to him which this brainless
bird cannot perform if expressly excited thereto; only the inner promptings
seem deficient, and when left to himself he spends most of his time crouched
on the ground with his head sunk between his shoulders as if asleep.[p.20]
GENERAL NOTION OF HEMISPHERES.
All these facts lead us, when we think about them, to some such explanatory
conception as this: The lower centres act from present sensational
stimuli alone; the hemispheres act from perceptions and considerations,
the sensations which they may receive, serving only as suggesters of these.
But what are perceptions but sensations grouped together? and what are
considerations but expectations, in the fancy, of sensations which will
be felt one way or another according as action takes this course or that?
If I step aside on seeing a rattlesnake, from considering how dangerous
an animal he is, the mental materials which constitute my prudential reflection
are images more or less vivid of the movement of his head, of a sudden
pain in my leg, of a state of terror, a swelling of the limb, a chill,
delirium, unconsciousness, etc., etc., and the ruin of my hopes. But all
these images are constructed out of my past experiences. They are reproductions
of what I have felt or witnessed. They are, in short, remote sensations;
and the difference between the hemisphereless animal and the whole one
may be concisely expressed by saying that the one obeys absent, the
other only present, objects.
The hemispheres would then seem to be the seat of memory. Vestiges
of past experience must in some way be stored up in them, and must, when
aroused by present stimuli, first appear as representations of distant
goods and evils; and then must discharge into the appropriate motor channels
for warding off the evil and securing the benefits of the good. If we liken
the nervous currents to electric currents, we can compare the nervous system,
C, below the hemispheres to a direct circuit from sense-organ to
muscle along the line S...C...M of Fig. 2 (p. 21). The hemisphere,
H, adds the long circuit or loop-line through which the current
may pass when for any reason the direct line is not used.
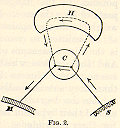 Thus, a tired wayfarer
on a hot day throws himself on [p.21] the damp earth beneath a maple-tree.
The sensations of delicious rest and coolness pouring themselves through
the direct line would naturally discharge into the muscles of complete
extension: he would abandon himself to the dangerous repose. But the loop-line
being open, part of the current is drafted along it, and awakens rheumatic
or catarral reminiscences, which prevail over the instigations of sense,
and make the man arise and pursue his way to where he may enjoy his rest
more safely. Presently we shall examine the manner in which the hemispheric
loop-line may be supposed to serve as a reservoir for such reminiscences
as these. Meanwhile I will ask the reader to notice some corollaries of
its being such a reservoir.
Thus, a tired wayfarer
on a hot day throws himself on [p.21] the damp earth beneath a maple-tree.
The sensations of delicious rest and coolness pouring themselves through
the direct line would naturally discharge into the muscles of complete
extension: he would abandon himself to the dangerous repose. But the loop-line
being open, part of the current is drafted along it, and awakens rheumatic
or catarral reminiscences, which prevail over the instigations of sense,
and make the man arise and pursue his way to where he may enjoy his rest
more safely. Presently we shall examine the manner in which the hemispheric
loop-line may be supposed to serve as a reservoir for such reminiscences
as these. Meanwhile I will ask the reader to notice some corollaries of
its being such a reservoir.
First, no animal without it can deliberate, pause, postpone, nicely
weigh one motive against another, or compare. Prudence, in a word, is for
such a creature an impossible virtue. Accordingly we see that nature removes
those functions in the exercise of which prudence is a virtue from the
lower centres and hands them over to the cerebrum. Wherever a creature
has to deal with complex features of the environment, prudence is a virtue.
The higher animals have so to deal; and the more complex the features,
the higher we call the animals. The fewer of his acts, then, can such
an animal perform without the help of the organs in question. In the frog
many acts devolve wholly on the lower centres; in the bird fewer; in the
rodent fewer still; in the dog very few indeed; and in apes and men hardly
any at all.
The advantages of this are obvious. Take the prehension of food as an
example and suppose it to be a reflex performance of the lower centres.
The animal will be condemned fatally and irresistibly to snap at it whenever
presented, no matter what the circumstances may be; he can no more disobey
this prompting than water can refuse to boil when a fire is kindled under
the pot. His life will again and again pay the forfeit of his gluttony.
[p.22] Exposure to retaliation, to other enemies, to traps, to poisons,
to the dangers of repletion, must be regular parts of his existence. His
lack of all thought by which to weigh the danger against the attractiveness
of the bait, and of all volition to remain hungry a little while longer,
is the direct measure of his lowness in the mental scale. And those fishes
which, like our cunners and sculpins, are no sooner thrown back from the
hook into the water, than they automatically seize the hook again, would
soon expiate the degradation of their intelligence by the extinction of
their type, did not their exaggerated fecundity atone for their imprudence.
Appetite and the acts it prompts have consequently become in all higher
vertebrates functions of the cerebrum. They disappear when the physiologist's
knife has left the subordinate centres alone in place. The brainless pigeon
will starve though left on a corn-heap.
Take again the sexual function. In birds this devolves exclusively upon
the hemispheres. When these are shorn away the pigeon pays no attention
to the billings and cooings of its mate. And Goltz found that a bitch in
heat would excite no emotion in male dogs who had suffered large loss of
cerebral tissue. Those who have read Darwin's 'Descent of Man' know what
immense importance in the amelioration of the breed in birds this author
ascribes to the mere fact of sexual selection. The sexual act is not performed
until every condition of circumstance and sentiment is fulfilled, until
time, place, and partner all are fit. But in frogs and toads this passion
devolves on the lower centres. They show consequently a machine-like obedience
to the present incitement of sense, and an almost total exclusion of the
power of choice. Copulation occurs per fas aut nefas, occasionally
between males, often with dead females, in puddles exposed on the highway,
and the male may be cut in two without letting go his hold. Every spring
an immense sacrifice of batrachian life takes place from these causes alone.
No one need be told how dependent all human social elevation is upon
the prevalence of chastity. Hardly any factor measures more than this the
difference between civili-[p.23] zation and barbarism. Physiologically
interpreted, chastity means nothing more than the fact that present solicitations
of sense are overpowered by suggestions of aesthetic and moral fitness
which the circumstances awaken in the cerebrum ; and that upon the inhibitory
or permissive influence of these alone action directly depends.
Within the psychic life due to the cerebrum itself the same general
distinction obtains, between considerations of the more immediate and considerations
of the more remote. In all ages the man whose determinations are swayed
by reference to the most distant ends has been held to possess the highest
intelligence. The tramp who lives from hour to hour; the bohemian whose
engagements are from day to day; the bachelor who builds but for a single
life; the father who acts for another generation ; the patriot who thinks
of a whole community and many generations; and finally, the philosopher
and saint whose cares are for humanity and for eternity,-these range themselves
in an unbroken hierarchy, wherein each successive grade results from an
increased manifestation of the special form of action by which the cerebral
centres are distinguished from all below them.
In the 'loop-line' along which the memories and ideas of the distant
are supposed to lie, the action, so far as it is a physical process, must
be interpreted after the type of the action in the lower centres. If regarded
here as a reflex process, it must be reflex there as well. The current
in both places runs out into the muscles only after it has first run in;
but whilst the path by which it runs out is determined in the lower centres
by reflections few and fixed amongst the cell-arrangements, in the hemispheres
the reflections are many and instable. This, it will be seen, is only a
difference of degree and not of kind, and does not change the reflex type.
The conception of all action as conforming to this type is the fundamental
conception of modern nerve-physiology. So much for our general preliminary
conception of the nerve-centres! Let us define it more distinctly before
we see how well physiological observation will bear it out in detail. [p.24]
THE EDUCATION OF THE HEMISPHERES
Nerve-currents run in through sense-organs, and whilst provoking reflex
acts in the lower centres, they arouse ideas in the hemispheres, which
either permit the reflexes in question, check them, or substitute others
for them. All ideas being in the last resort reminiscences, the question
to answer is: How can processes become organized in the hemispheres
which correspond to reminiscences in the mind ?[3]
Nothing is easier than to conceive a possible way in which this might
be done, provided four assumptions be granted. These assumptions (which
after all are inevitable in any event) are:
1) The same cerebral process which, when aroused from without by a sense-organ,
gives the perception of an object, will give an idea of the same
object when aroused by other cerebral processes from within.
2) If processes 1, 2, 3, 4 have once been aroused together or in immediate
succession, any subsequent arousal of any one of them (whether from without
or within) will tend to arouse the others in the original order.[This
is the so-called law of association.]
3) Every sensorial excitement propagated to a lower centre tends to
spread upwards and arouse an idea.
4) Every idea tends ultimately either to produce a movement or to check
one which otherwise would be produced.
Suppose now (these assumptions being granted) that we have a baby before
us who sees a candle-flame for the first [p. 25] time, and, by virtue of
a reflex tendency common in babies of a certain age, extends his hand to
grasp it, so that his fingers get burned. So far we have two reflex currents
in play: first, from the eye to the extension movement, along the line
1-1-1-1 of Fig. 3; and second, from the finger to the movement of drawing
back the hand, along the line 2-2-2-2.
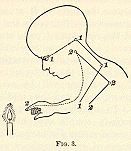 If this were the baby's whole nervous
system, and if the reflexes were once for all organic, we should have no
alteration in his behavior, no matter how often the experience recurred.
The retinal image of the flame would always make the arm shoot forward,
the burning of the finger would always send it back. But we know that 'the
burnt child dreads the fire,' and that one experience usually protects
the fingers forever. The point is to see how the hemispheres may bring
this result to pass.
If this were the baby's whole nervous
system, and if the reflexes were once for all organic, we should have no
alteration in his behavior, no matter how often the experience recurred.
The retinal image of the flame would always make the arm shoot forward,
the burning of the finger would always send it back. But we know that 'the
burnt child dreads the fire,' and that one experience usually protects
the fingers forever. The point is to see how the hemispheres may bring
this result to pass.
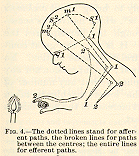 We must complicate our diagram (see
Fig. 4). Let the current 1-1, from the eye, discharge upward as well as
downward when it reaches the lower centre for vision, and arouse the perceptional
process s1 in the hemispheres; let the feeling of the
arm's extension also send up a current which leaves a trace of itself,
m1; let the burnt finger leave an analogous trace, s2;
and let the movement of retraction leave m2. These four
processes will now, by virtue of assumption 2), be associated together
by the path s1-m1-s2-m2
running from the first to the last, so that if anything touches off s1,
ideas of the extension, of the burnt finger, and of the retraction will
pass in rapid succession [p.26] through the mind. The effect on the child's
conduct when the candle-flame is next presented is easy to imagine. Of
course the sight of it arouses the grasping reflex; but it arouses simultaneously
the idea thereof, together with that of the consequent pain, and of the
final retraction of the hand; and if these cerebral processes prevail in
strength over the immediate sensation in the centres below, the last idea
will be the cue by which the final action is discharged. The grasping will
be arrested in mid-career, the hand drawn back, and the child's fingers
saved.
We must complicate our diagram (see
Fig. 4). Let the current 1-1, from the eye, discharge upward as well as
downward when it reaches the lower centre for vision, and arouse the perceptional
process s1 in the hemispheres; let the feeling of the
arm's extension also send up a current which leaves a trace of itself,
m1; let the burnt finger leave an analogous trace, s2;
and let the movement of retraction leave m2. These four
processes will now, by virtue of assumption 2), be associated together
by the path s1-m1-s2-m2
running from the first to the last, so that if anything touches off s1,
ideas of the extension, of the burnt finger, and of the retraction will
pass in rapid succession [p.26] through the mind. The effect on the child's
conduct when the candle-flame is next presented is easy to imagine. Of
course the sight of it arouses the grasping reflex; but it arouses simultaneously
the idea thereof, together with that of the consequent pain, and of the
final retraction of the hand; and if these cerebral processes prevail in
strength over the immediate sensation in the centres below, the last idea
will be the cue by which the final action is discharged. The grasping will
be arrested in mid-career, the hand drawn back, and the child's fingers
saved.
In all this we assume that the hemispheres do not natively couple
any particular sense-impression with any special motor discharge. They
only register, and preserve traces of, such couplings as are already organized
in the reflex centres below. But this brings it inevitably about that,
when a chain of experiences has been already registered and the first link
is impressed once again from without, the last link will often be awakened
in idea long before it can exist in fact. And if this last
link were previously coupled with a motion, that motion may now come from
the mere ideal suggestion without waiting for the actual impression to
arise. Thus an animal with hemispheres acts in anticipation of future
things; or, to use our previous formula, he acts from considerations of
distant good and ill. If we give the name of partners to the original
couplings of impressions with motions in a reflex way, then we may say
that the function of the hemispheres is simply to bring about exchanges
among the partners. Movement mn, which natively is
sensation sn's partner, becomes through the hemispheres
the partner of sensation s1, s2 or
s3. It is like the great commutating switch-board at
a central telephone station. No new elementary process is involved; no
impression nor any motion peculiar to the hemispheres; but any number of
combinations impossible to the lower machinery taken alone, and an endless
consequent increase in the possibilities of behavior on the creature's
part.
All this, as a mere scheme,[4] is so clear and so
concordant [p.27] with the general look of the facts as almost to impose
itself on our belief; but it is anything but clear in detail. The brain-physiology
of late years has with great effort sought to work out the paths by which
these couplings of sensations with movements take place, both in the hemispheres
and in the centres below.
So we must next test our scheme by the facts discovered in this direction.
We shall conclude, I think, after taking them all into account, that the
scheme probably makes the lower centres too machine-like and the hemispheres
not quite machine-like enough, and must consequently be softened down a
little. So much I may say in advance. Meanwhile, before plunging into the
details which await us, it will somewhat clear our ideas if we contrast
the modern way of looking at the matter with the phrenological conception
which but lately preceded it.
THE PHRENOLOGICAL CONCEPTION.
In a certain sense Gall was the first to seek to explain in detail how
the brain could subserve our mental operations. His way of proceeding was
only too simple. He took the faculty-psychology as his ultimatum on the
mental side, and he made no farther psychological analysis. Wherever
he found an individual with some strongly-marked trait of character he
examined his head; and if he found the latter prominent in a certain region,
he said without more ado that that region was the 'organ' of the trait
or faculty in question. The traits were of very diverse constitution, some
being simple sensibilities like 'weight' or 'color'; some being instinctive
tendencies like 'alimentiveness' or 'amativeness;' and others, again, being
complex resultants like 'conscientiousness,' 'individuality.' Phrenology
fell promptly into disrepute among scientific men because observation seemed
to show that large facul-[p.28] ties and large 'bumps' might fail to coexist;
because the scheme of Gall was so vast as hardly to admit of accurate determination
at all-who of us can say even of his own brothers whether their perceptions
of weight and of time are well developed or not?-because
the followers of Gall and Spurzheim were unable to reform these errors
in any appreciable degree; and, finally, because the whole analysis of
faculties was vague and erroneous from a psychologic point of view. Popular
professors of the lore have nevertheless continued to command the admiration
of popular audiences; and there seems no doubt that Phrenology, however
little it satisfy our scientific curiosity about the functions of different
portions of the brain, may still be, in the hands of intelligent practitioners,
a useful help in the art of reading character. A hooked nose and a firm
jaw are usually signs of practical energy; soft, delicate hands are signs
of refined sensibility. Even so may a prominent eye be a sign of power
over language, and a bull-neck a sign of sensuality. But the brain behind
the eye and neck need no more be the organ of the signified faculty
than the jaw is the organ of the will or the hand the organ of refinement.
These correlations between mind and body are, however, so frequent that
the 'characters' given by phrenologists are often remarkable for knowingness
and insight.
Phrenology hardly does more than restate the problem. To answer the
question, "Why do I like children?" by saying, "Because you have a large
organ of philoprogenitiveness," but renames the phenomenon to be explained.
What is my philoprogenitiveness? Of what mental elements does it
consist? And how can a part of the brain be its organ? A science
of the mind must reduce such complex manifestations as 'philoprogenitiveness'
to their elements. A science of the brain must point out the functions
of its elements. A science of the relations of mind and brain must
show how the elementary ingredients of the former correspond to the elementary
functions of the latter. But phrenology, except by occasional coincidence,
takes no account of elements at all. Its 'faculties,' as a rule, are fully
equipped persons in a particular mental attitude. Take, for example, the
'faculty' of language. It involves [p.29] in reality a host of distinct
powers. We must first have images of concrete things and ideas of abstract
qualities and relations; we must next have the memory of words and then
the capacity so to associate each idea or image with a particular word
that, when the word is heard, the idea shall forthwith enter our mind.
We must conversely, as soon as the idea arises in our mind, associate with
it a mental image of the word, and by means of this image we must innervate
our articulatory apparatus so as to reproduce the word as physical sound.
To read or to write a language other elements still must be introduced.
But it is plain that the faculty of spoken language alone is so complicated
as to call into play almost all the elementary powers which the mind possesses,
memory, imagination, association, judgment, and volition. A portion of
the brain competent to be the adequate seat of such a faculty would needs
be an entire brain in miniature,-just as the faculty itself is really a
specification of the entire man, a sort of homunculus. Yet just such homunculi
are for the most part the phrenological organs. As Lange says:
"We have a parliament of little men together, each of whom, as happens
also in a real parliament, possesses but a single idea which he ceaselessly
strives to make prevail"-benevolence, firmness, hope, and the rest. "Instead
of one soul, phrenology gives us forty, each alone as enigmatic as the
full aggregate psychic life can be. Instead of dividing the latter into
effective elements, she divides it into personal beings of peculiar character
..'Herr Pastor, sure there be a horse inside,' called out the peasants
to X after their spiritual shepherd had spent hours in explaining to them
the construction of the locomotive. With a horse inside truly everything
becomes clear, even though it be a queer enough sort of horse-the horse
itself calls for no explanation! Phrenology takes a start to get beyond
the point of view of the ghost-like soul entity, but she ends by populating
the whole skull with ghosts of the same order."[5]
Modern Science conceives of the matter in a very different way. Brain
and mind alike consist of simple elements, sensory and motor.
"All nervous centres," says Dr. Hughlings Jackson,[6]
"from the lowest to the very highest (the [p.30] substrata of consciousness),
are made up of nothing else than nervous arrangements, representing impressions
and movements. . . I do not see of what other materials the brain can
be made." Meynert represents the matter similarly when he calls the cortex
of the hemispheres the surface of projection for every muscle and every
sensitive point of the body. The muscles and the sensitive points are represented
each by a cortical point, and the brain is nothing but the sum of all these
cortical points, to which, on the mental side, as many ideas correspond.
Ideas of sensation, ideas of motion are, on the other hand, the
elementary factors out of which the mind is built up by the associationists
in psychology. There is a complete parallelism between the two analyses,
the same diagram of little dots, circles, or triangles joined by lines
symbolizes equally well the cerebral and mental processes : the dots stand
for cells or ideas, the lines for fibres or associations. We shall have
later to criticise this analysis so far as it relates to the mind; but
there is no doubt that it is a most convenient, and has been a most useful,
hypothesis, formulating the facts in an extremely natural way.
If, then, we grant that motor and sensory ideas variously associated
are the materials of the mind, all we need do to get a complete diagram
of the mind's and the brain's relations should be to ascertain which sensory
idea corresponds to which sensational surface of projection, and which
motor idea to which muscular surface of projection. The associations would
then correspond to the fibrous connections between the various surfaces.
This distinct cerebral localization of the various elementary
sorts of idea has been treated as a 'postulate' by many physiologists (e.g.
Munk); and the most stirring controversy in nerve-physiology which the
present generation has seen has been the localization-question.
THE LOCALIZATION OF FUNCTIONS IN THE.
HEMISPHERES.
Up to 1870, the opinion which prevailed was that which the experiments
of Flourens on pigeons' brains had made plausible, namely, that the different
functions of the hemi-[p.31] spheres were not locally separated, but carried
on each by the aid of the whole organ. Hitzig in 1870 showed, however,
that in a dog's brain highly specialized movements could be produced by
electric irritation of determinate regions of the cortex; and Ferrier and
Munk, half a dozen years later, seemed to prove, either by irritations
or excisions or both, that there were equally determinate regions connected
with the senses of sight, touch, hearing, and smell. Munk's special sensorial
localizations, however, disagreed with Ferrier's; and Goltz, from his extirpation-experiments,
came to a conclusion adverse to strict localization of any kind. The controversy
is not yet over. I will not pretend to say anything more of it historically,
but give a brief account of the condition in which matters at present stand.
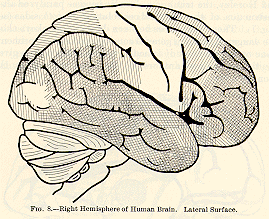
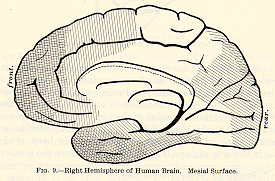
The one thing which is perfectly well established is this, that
the 'central' convolutions, on either side of the fissure of Rolando, and
(at least in the monkey) the calloso-marginal convolution (which is continuous
with them on the mesial surface where one hemisphere is applied against
the other), form the region by which all the motor incitations which leave
the cortex pass out, on their way to those executive centres in the region
of the pons, medulla, and spinal cord from which the muscular contractions
are discharged in the last resort. The existence of this so-called 'motor
zone' is established by the lines of evidence successively given below:
(1) Cortical Irritations. Electrical currents of small intensity
applied to the surface of the said convolutions in dogs, monkeys, and other
animals, produce well-defined movements in face, fore-limb, hind-limb,
tail, or trunk, according as one point or another of the surface is irritated.
These movements affect almost invariably the side opposite to the brain
irritations : If the left hemisphere be excited, the movement is of the
right leg, side of face, etc. All the objections at first raised against
the validity of these experiments have been overcome. The movements are
certainly not due to irritations of the base of the brain by the downward
spread of the current, for: a) mechanical irritations will produce
them, though less easily than electrical; b) shifting the [p.32]
electrodes to a point close by on the surface changes the movement in ways
quite inexplicable by changed physical conduction of the current;
c) if the cortical 'centre' for a certain movement be cut under
with a sharp knife but left in situ, although the electric conductivity
is physically unaltered by the operation, the physiological conductivity
is gone and currents of the same strength no longer produce the movements
which they did; d) the time-interval between the application of
the electric stimulus to the cortex and the resultant movement is what
it would be if the cortex acted physiologically and not merely physically
in transmitting the irritation. It is namely a well-known fact that when
a nerve-current has to pass through the spinal cord to excite a
muscle by reflex action, the time is longer than if it passes directly
down the motor nerve: the cells of the cord take a certain time to discharge.
Similarly, when a stimulus is applied directly to the cortex the muscle
contracts two or three hundredths of a second later than it does when the
place on the cortex is cut away and the electrodes are applied to the white
fibres below.[7]
(2) Cortical Ablations. When the cortical spot which is found
to produce a movement of the fore-leg, in a dog, is excised (see
spot 5 in Fig. 5), the leg in question becomes peculiarly affected. At
first it seems paralyzed. Soon, however, it is used with the other legs,
but badly. The animal does not bear his weight on it, allows it to rest
on its dorsal surface, stands with it crossing the other leg, does not
remove it if it hangs over the edge of a table, can no longer 'give
the paw' at word of command if able to do so before the operation, does
not use it for scratching the ground, or holding a bone as formerly, lets
it slip out when running on a smooth [p.33] surface or when shaking himself,
etc., etc. Sensibility of all kinds seems diminished as well as motility,
but of this I shall speak later on. Moreover the dog tends in voluntary
movements to swerve towards the side of the brain-lesion instead of going
straight forward. All these symptoms gradually decrease, so that even with
a very severe brain-lesion the dog may be outwardly indistinguishable from
a well dog after eight or ten weeks. Still, a slight chloroformization
will reproduce the disturbances, even then. There is a certain appearance
of ataxic in-coördination in the movements -the dog lifts his fore-feet
high and brings them down with more strength than usual, and yet the trouble
is not ordinary lack of co-ordination.
Neither is there paralysis. The strength of whatever movements are made
is as great as ever-dogs with extensive destruction of the motor zone can
jump as high and bite as hard as ever they did, but they seem less easily
moved to do anything with the affected parts. Dr. Loeb, who
has studied the motor disturbances of dogs more carefully than any one,
conceives of them en masse as effects of an increased inertia in
all the processes of innervation towards the side opposed to the lesion.
All such movements require an unwonted effort for their execution; and
when only the normally usual effort is made they fall behind in effectiveness.[8]
[p.34] Even when the entire motor zone of a dog is removed, there is
no permanent paralysis of any part, but only this curious sort of relative
inertia when the two sides of the body are compared; and this itself becomes
hardly noticeable after a number of weeks have elapsed. Prof Goltz has
described a dog whose entire left hemisphere was destroyed, and who retained
only a slight motor inertia on the right half of the body. In particular
he could use his right paw for holding a bone whilst gnawing it, or for
reaching after a piece of meat.
Had he been taught to give his paw before the operations, it would have
been curious to see whether that faculty also came back. His tactile sensibility
was permanently diminished on the right side.[9] In monkeys
a genuine paralysis follows upon ablations of the cortex in the motor region.
This paralysis affects parts of the body which vary with the brain-parts
removed. The monkey's opposite arm or leg hangs flaccid, or at most takes
a small part in associated movements. When the entire region is removed
there is a genuine and permanent hemiplegia in which the arm is more affected
than the leg; and this is [p.35] followed months later by contracture of
the muscles, as in man after inveterate hemiplegia.[10]
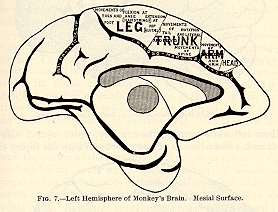
According to Schaefer and Horsley, the trunk-muscles also become paralyzed
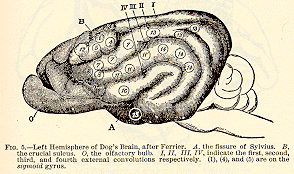
after destruction of the marginal convolution on both sides
(see Fig. 7). These differences between dogs and monkeys show the danger
of drawing general conclusions from experiments done on any one sort of
animal. I subjoin the figures given by the last-named authors of the motor
regions in the monkey's brain.[11]
In man we are necessarily reduced to the observation post-mortem
of cortical ablations produced by accident or disease (tumor, hemorrhage,
softening, etc.). What results during life from such conditions is either
localized spasm, or palsy of certain muscles of the opposite side. The
cortical regions which invariably produce these results are homologous
with those which we have just been studying in the dog, cat, ape, etc.
Figs. 8 and 9 show the result of [p.36] 169 cases carefully studied by
Exner. The parts shaded are regions where lesions produced no motor
disturbance. Those left white were, on the contrary, never injured without
motor disturbances of some sort.
Where the injury to the cortical substance is profound in man, the paralysis
is permanent and is succeeded by muscular rigidity in the paralyzed parts,
just as it may be in the monkey. [p.37]
(3) Descending degenerations show the intimate connection of
the rolandic regions of the cortex with the motor tracts of the cord. When,
either in man or in the lower animals, these regions are destroyed, a peculiar
degenerative change known as secondary sclerosis is found to extend downwards
through the white fibrous substance of the brain in a perfectly definite
manner, affecting certain distinct strands which pass through the inner
capsule, crura, and pons, into the anterior pyramids of the medulla oblongata,
and from thence (partly crossing to the other side) downwards into the
anterior (direct) and lateral (crossed) columns of the spinal cord.
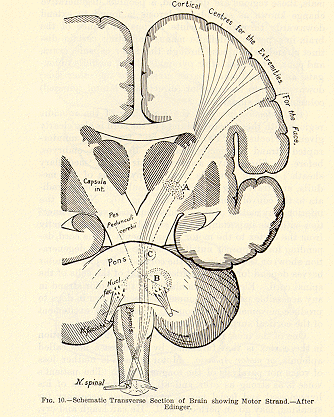
(4) Anatomical proof of the continuity of the rolandic regions
with these motor columns of the cord is also clearly given. Flechsig's
'Pyramidenbahn' forms an uninterrupted strand (distinctly traceable in
human embryos, before its fibres have acquired their white 'medullary sheath')
passing upwards from the pyramids of the medulla, and traversing the internal
capsule and corona radiata to the convolutions in question (Fig. 10). None
of the inferior gray matter of the brain seems to have any connection with
this important fibrous strand. It passes directly from the cortex to the
motor arrangements in the cord, depending for its proper nutrition (as
the facts of degeneration show) on the influence of the cortical cells,
just as motor nerves depend for their nutrition on that of the cells of
the spinal cord. Electrical stimulation of this motor strand in any accessible
part of its course has been shown in dogs to produce movements analogous
to those which excitement of the cortical surface calls forth.
One of the most instructive proofs of motor localization in the cortex
is that furnished by the disease now called aphemia, or motor Aphasia.
Motor aphasia is neither loss of voice nor paralysis of the tongue or lips.
The patient's voice is as strong as ever, and all the innervations of his
hypoglossal and facial nerves, except those necessary for speaking, may
go on perfectly well. He can laugh and cry, and even sing; but he either
is unable to utter any words at all; or a few meaningless stock phrases
form his only speech ; or else he speaks incoherently and confusedly, mispronounc-[p.38]
ing, misplacing, and misusing his words in various degrees. Sometimes his
speech is a mere broth of unintelligible syllables. In cases of pure motor
aphasia the patient recognizes his mistakes and suffers acutely from them.
Now whenever a patient dies in such a condition as this, and an examination
of his brain is permitted, it is found that [p.39] the lowest frontal gyrus
(see Fig. 11) is the seat of injury. Broca first noticed this fact in 1861,
and since then the gyrus has gone by the name of Broca's convolution.
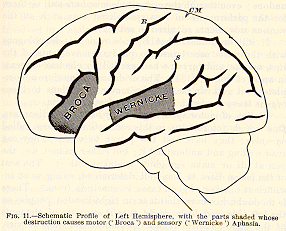
The injury in right-handed people is found on the left hemisphere, and
in left-handed people on the right hemisphere. Most people, in fact, are
left-brained, that is, all their delicate and specialized movements are
handed over to the charge of the left hemisphere. The ordinary right-handedness
for such movements is only a consequence of that fact, a consequence which
shows outwardly on account of that extensive decussation of the fibres
whereby most of those from the left hemisphere pass to the right half of
the body only. But the left-brainedness might exist in equal measure and
not show outwardly. This would happen wherever organs on both sides
of the body could be governed by the left hemisphere; and just such a case
seems offered by the vocal organs, in that highly delicate and special
motor service which we call speech. Either hemisphere can innervate
them bilaterally, just as either seems able to innervate bilaterally the
muscles of the trunk, ribs, and diaphragm. Of the special movements of
speech, how-[p.40] ever, it would appear (from the facts of aphasia) that
the left hemisphere in most persons habitually takes exclusive charge.
With that hemisphere thrown out of gear, speech is undone; even though
the opposite hemisphere still be there for the performance of less specialized
acts, such as the various movements required in eating.
It will be noticed that Broca's region is homologous with the parts
ascertained to produce movements of the lips, tongue, and larynx when excited
by electric currents in apes (cf. Fig. 6, p. 34). The evidence is therefore
as complete as it well can be that the motor incitations to these organs
leave the brain by the lower frontal region.
Victims of motor aphasia generally have other disorders. One which interests
us in this connection has been called agraphia: they have lost the
power to write. They can read writing and understand it; but either
cannot use the pen at all or make egregious mistakes with it. The seat
of the lesion here is less well determined, owing to an insufficient number
of good cases to conclude from.[12] There is no doubt,
however, that it is (in right-handed people) on the left side, and little
doubt that it consists of elements of the hand-and-arm region specialized
for that service. The symptom may exist when there is little or no disability
in the hand for other uses. If it does not get well, the patient usually
educates his right hemisphere, i.e. learns to write with his left hand.
In other cases of which we shall say more a few pages later on, the patient
can write both spontaneously and at dictation, but cannot read even
what he has himself written! All these phenomena are now quite clearly
explained by separate brain-centres for the various feelings and movements
and tracts for associating these together. But their minute discussion
belongs to medicine rather than to general psychology, and I can only use
them here to illustrate the principles of motor localization.[13]
Under the heads of sight and hearing I shall have a little more to say.
[p.41] The different lines of proof which I have taken up establish
conclusively the proposition that all the motor impulses which
leave the cortex pass out, in healthy animals, from the convolutions
about the fissure of Rolando.
When, however, it comes to defining precisely what is involved in a
motor impulse leaving the cortex, things grow more obscure. Does the impulse
start independently from the convolutions in question, or does it start
elsewhere and merely flow through? And to what particular phase of psychic
activity does the activity of these centres correspond? Opinions and authorities
here divide; but it will be better, before entering into these deeper aspects
of the problem, to cast a glance at the facts which have been made out
concerning the relations of the cortex to sight, hearing, and smell.
Sight.
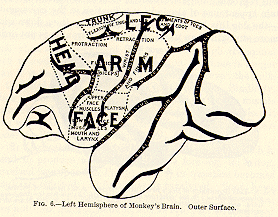
Ferrier was the first in the field here. He found, when the angular
convolution (that lying between the 'intra parietal' and 'external occipital'
fissures, and bending round the top of the fissure of Sylvius, in Fig.
6) was excited in the monkey, that movements of the eyes and head as if
for vision occurred; and that when it was extirpated, what he supposed
to be total and permanent blindness of the opposite eye followed. Munk
almost immediately declared total and permanent blindness to follow from
destruction of the occipital lobe in monkeys as well as dogs, and
said that the angular gyrus had nothing to do with sight, but was only
the centre for tactile sensibility of the eyeball. Munk's absolute tone
about his observations and his theoretic arrogance have led to his ruin
as an authority. But he did two things of permanent value. He was the first
to distinguish in these vivisections between sensorial and psychic
blindness, and to describe the phenomenon of restitution of the
visual function after its first impairment by an operation; and the first
to notice the hemiopic character of the visual disturbances which
result when only one hemisphere is injured. Sensorial blindness is absolute
insensibility to light; psychic blindness is inability to recognize the
meaning of the optical impressions, as when we [p.42] see a page
of Chinese print but it suggests nothing to us. A hemiopic disturbance
of vision is one in which neither retina is affected in its totality, but
in which, for example, the left portion of each retina is blind,
so that the animal sees nothing situated in space towards its right. Later
observations have corroborated this hemiopic character of all the disturbances
of sight from injury to a single hemisphere in the higher animals; and
the question whether an animal's apparent blindness is sensorial
or only psychic has, since Munk's first publications, been the most urgent
one to answer, in all observations relative to the function of sight.
Goltz almost simultaneously with Ferrier and Munk reported experiments
which led him to deny that the visual function was essentially bound up
with any one localized portion of the hemispheres. Other divergent results
soon came in from many quarters, so that, without going into the history
of the matter any more, I may report the existing state of the case as
follows:[14]
In fishes, frogs, and lizards vision persists when the
hemispheres are entirely removed. This is admitted for frogs and fishes
even by Munk, who denies it for birds.
All of Munk's birds seemed totally blind (blind sensorially)
after removal of the hemispheres by his operation. The following of a candle
by the head and winking at a threatened blow, which are ordinarily held
to prove the retention of crude optical sensations by the lower centres
in supposed hemisphereless pigeons, are by Munk ascribed to vestiges of
the visual sphere of the cortex left behind by the imperfection of the
operation. But Schrader, who operated after Munk and with every apparent
guarantee of completeness, found that all his pigeons saw after
two or three weeks had elapsed, and the inhibitions resulting from
the wound had passed away. They invariably avoided even the slightest obstacles,
flew very regularly towards certain perches, etc., differing toto coelo
in these respects with certain simply blinded pigeons who were kept
with [p.43] them for comparison. They did not pick up food strewn on the
ground, however. Schrader found that they would do this if even a small
part of the frontal region of the hemispheres was left, and ascribes their
non-self-feeding when deprived of their occipital cerebrum not to a visual,
but to a motor, defect, a sort of alimentary aphasia.[15]
In presence of such discord as that between Munk and his opponents one
must carefully note how differently significant is loss, from preservation,
of a function after an operation on the brain. The loss of the function
does not necessarily show that it is dependent on the part cut out; but
its preservation does show that it is not dependent: and this is
true though the loss should be observed ninety-nine times and the preservation
only once in a hundred similar excisions. That birds and mammals can
be blinded by cortical ablation is undoubted; the only question is, must
they be so? Only then can the cortex be certainly called the 'seat of sight.'
The blindness may always be due to one of those remote effects of the wound
on distant parts, inhibitions, extensions of inflammation,-interferences,
in a word,- upon which Brown-Séquard and Goltz have rightly insisted,
and the importance of which becomes more manifest every day. Such effects
are transient; whereas the symptoms of deprivation (Ausfallserscheinungen,
as Goltz calls them) which come from the actual loss of the cut-out region
must from the nature of the case be permanent. Blindness in the pigeons,
so far as it passes away, cannot possibly be charged to their seat
of vision being lost, but only to some influence which temporarily depresses
the activity of that seat. The same is true mutatis mutandis of
all the other effects of operations, and as we pass to mammals we shall
see still more the importance of the remark.
In rabbits loss of the entire cortex seems compatible with the
preservation of enough sight to guide the poor animals' movements, and
enable them to avoid obstacles. Christiani's observations and discussions
seem conclusively [p.44] to have established this, although Munk found
that all his animals were made totally blind.[16]
In dogs also Munk found absolute stone-blindness after ablation
of the occipital lobes. He went farther and mapped out determinate portions
of the cortex thereupon, which he considered correlated with definite segments
of the two retinae, so that destruction of given portions of the cortex
produces blindness of the retinal centre, top, bottom, or right or left
side, of the same or opposite eye. There seems little doubt that this definite
correlation is mythological. Other observers, Hitzig, Goltz, Luciani, Loeb,
Exner, etc., find, whatever part of the cortex may be ablated on one side,
that there usually results a hemiopic disturbance of both
eyes, slight and transient when the anterior lobes are the parts attacked,
grave when an occipital lobe is the seat of injury, and lasting in proportion
to the latter's extent. According to Loeb, the defect is a dimness of vision
('hemiamblyopia') in which (however severe) the centres remain the best
seeing portions of the retina, just as they are in normal dogs. The lateral
or temporal part of each retina seems to be in exclusive connection with
the cortex of its own side. The centre and nasal part of each seems, on
the contrary, to be connected with the cortex of the opposite hemispheres.
Loeb, who takes broader views than any one, conceives the hemiamblyopia
as he conceives the motor disturbances, namely, as the expression of an
increased inertia in the whole optical machinery, of which the result is
to make the animal respond with greater effort to impressions coming from
the half of space opposed to the side of the lesion. If a dog has right
hemiamblyopia, say, and two pieces of meat are hung before him at once,
he invariably turns first to the one on his left. But if the lesion be
a slight one, shaking slightly the piece of meat on his right (this
makes of it a stronger stimulus) makes him seize upon it first. If only
one piece of meat be offered, he takes it, on whichever side it be.
When both occipital lobes are extensively destroyed total blindness
may result. Munk maps out his 'Seh-[p.45] sphäre' definitely, and
says that blindness must result when the entire shaded part, marked A,
A, in Figs. 12 and 13, is involved in the lesion. Discrepant reports of
other observations he explains as due to incomplete ablation.
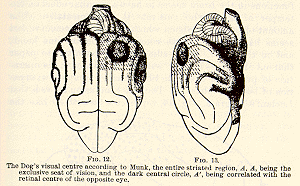
Luciani, Goltz, and Lannegrace, however, contend that they have made
complete bilateral extirpations of Munk's Sehsphäre more than once,
and found a sort of crude indiscriminating sight of objects to return in
a few weeks.[17] The question whether a dog is blind
or not is harder to solve than would at first appear; for simply blinded
dogs, in places to which they are accustomed, show little of their loss
and avoid all obstacles; whilst dogs whose occipital lobes are gone may
run against things frequently and yet see notwithstanding. The best proof
that they may see is that which Goltz's dogs furnished: they carefully
avoided, as it seemed, strips of sunshine or paper on the floor, as if
they were solid obstacles. This no really blind dog would do. Luciani tested
his dogs when hungry (a condition which sharpens their attention) by strewing
[p.46] pieces of meat and pieces of cork before them. If they went straight
at them, they saw; and if they chose the meat and left the cork,
they saw discriminatingly. The quarrel is very acrimonious; indeed
the subject of localization of functions in the brain seems to have a peculiar
effect on the temper of those who cultivate it experimentally. The amount
of preserved vision which Goltz and Luciani report seems hardly to be worth
considering, on the one hand; and on the other, Munk admits in his penultimate
paper that out of 85 dogs he only 'succeeded' 4 times in his operation
of producing complete blindness by complete extirpation of his 'Sehsphäre'.[18]
The safe conclusion for us is that Luciani's diagram, Fig. 14, represents
something like the truth.
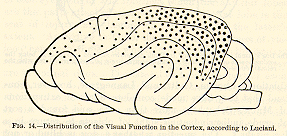
The occipital lobes are far more important for vision than any other
part of the cortex, so that their complete destruction makes the animal
almost blind. As for the crude sensibility to light which may then
remain, nothing exact is known either about its nature or its seat.
In the monkey, doctors also disagree. The truth seems, however,
to be that the occipital lobes in this animal also are the part
connected most intimately with the visual function. The function would
seem to go on when very small portions of them are left, for Ferrier found
no 'appreciable impairment' of it after almost complete destruction of
them on both sides. On the other hand, he found complete and permanent
blindness to ensue when they and the angular gyri in addition
were destroyed on both sides. Munk, as well as [p.47] Brown and Schaefer,
found no disturbance of sight from destroying the angular gyri alone,
although Ferrier found blindness to ensue. This blindness was probably
due to inhibitions exerted in distans, or to cutting of the white
optical fibres passing under the angular gyri on their way to the occipital
lobes. Brown and Schaefer got complete and permanent blindness in one monkey
from total destruction of both occipital lobes. Luciani and Seppili, performing
this operation on two monkeys, found that the animals were only mentally,
not sensorially, blind. After some weeks they saw their food, but could
not distinguish by sight between figs and pieces of cork. Luciani and Seppili
seem, however, not to have extirpated the entire lobes. When one lobe only
is injured the affection of sight is hemiopic in monkeys: in this all observers
agree. On the whole, then, Munk's original location of vision in the occipital
lobes is confirmed by the later evidence.[19]
In man we have more exact results, since we are not driven to
interpret the vision from the outward conduct. On the other hand, however,
we cannot vivisect, but must wait for pathological lesions to turn up.
The pathologists who have discussed these (the literature is tedious ad
libitum) conclude that the occipital lobes are the indispensable part
for vision in man. Hemiopic disturbance in both eyes comes from lesion
of either one of them, and total blindness, sensorial as well as psychic,
from destruction of both.
Hemiopia may also result from lesion in other parts, especially the
neighboring angular and supra-marginal gyri, and it may accompany extensive
injury in the motor region of the cortex. In these cases it seems probable
that it is due to an actio in distans, probably to the interruption
of [p.48] fibres proceeding from the occipital lobe. There seem to be a
few cases on record where there was injury to the occipital lobes without
visual defect. Ferrier has collected as many as possible to prove his localization
in the angular gyrus.[20] A strict application of logical
principles would make one of these cases outweigh one hundred contrary
ones. And yet, remembering how imperfect observations may be, and how individual
brains may vary, it would certainly be rash for their sake to throw away
the enormous amount of positive evidence for the occipital lobes. Individual
variability is always a possible explanation of an anomalous case.
There is no more prominent anatomical fact than that of the 'decussation
of the pyramids,' nor any more usual pathological fact than its consequence,
that left-handed hemorrhages into the motor region produce right-handed
paralyses. And yet the decussation is variable in amount, and seems sometimes
to be absent altogether.[21] If, in such a case as this
last, the left brain were to become the seat of apoplexy, the left
and not the right half of the body would be the one to suffer paralysis.
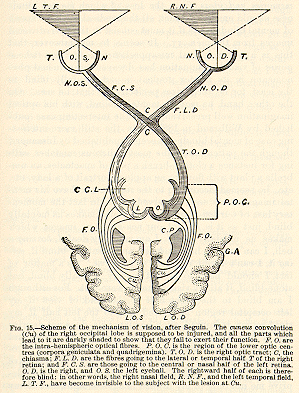
The schema on the opposite page, copied from Dr.Seguin, expresses,
on the whole, the probable truth about the regions concerned in vision.
Not the entire occipital lobes, but the so-called cunei, and the first
convolutions, are the cortical parts most intimately concerned. Nothnagel
agrees with Seguin in this limitation of the essential tracts.[22]
A most interesting effect of cortical disorder is mental blindness.
This consists not so much in insensibility to optical impressions, as in
inability to understand them. Psychologically it is interpretable
as loss of associations between optical sensations and what they
signify; and any interruption of the paths between the optic centres and
the centres for other ideas ought to bring it about. Thus,[p.49] printed
letters of the alphabet, or words, signify certain sounds and certain articulatory
movements. If the connection between the articulating or auditory centres,
on the one hand, and the visual centres on the other, be ruptured, we ought
a priori to expect that the sight of words would fail to awaken
the idea of their sound, or the movement for pronouncing them.
We ought, in short, to have alexia, or inability to read: and
this is just what we do have in many [p.50] cases of extensive injury about
the fronto-temporal regions, as a complication of aphasic disease.
Nothnagel suggests that whilst the cuneus is the seat of optical
sensations, the other parts of the occipital lobe may be the field
of optical memories and ideas, from the loss of which mental blindness
should ensue. In fact, all the medical authors speak of mental blindness
as if it must consist in the loss of visual images from the memory. It
seems to me, however, that this is a psychological misapprehension. A man
whose power of visual imagination has decayed (no unusual phenomenon in
its lighter grades) is not mentally blind in the least, for he recognizes
perfectly all that he sees. On the other hand, he may be mentally
blind, with his optical imagination well preserved; as in the interesting
case publislied by Wilbrand in 1887.[23] In the still
more interesting case of mental blindness recently published by Lissauer,[24]
though the patient made the most ludicrous mistakes, calling for instance
a clothes-brush a pair of spectacles, an umbrella a plant with flowers,
an apple a portrait of a lady, etc. etc., he seemed, according to the reporter,
to have his mental images fairly well preserved. It is in fact the momentary
loss of our non-optical images which makes us mentally blind, just
as it is that of our non-auditory images which makes us mentally
deaf. I am mentally deaf if, hearing a bell, I can't recall how
it looks; and mentally blind if, seeing it, I can't recall
its sound or its name. As a matter of fact, I should have to be
not merely mentally blind, but stone-blind, if all my visual images
were lost. For although I am blind to the right half of the field of view
if my left occipital region is injured, and to the left half if my right
region is injured, such hemianopsia does not deprive me of visual images,
experience seeming to show that the unaffected hemisphere is always sufficient
for production of these. To abolish them entirely I should have to be deprived
of both occipital lobes, and that would deprive me not only of my inward
images of sight, but of my [p.51] sight altogether.[25]
Recent pathological annals seem to offer a few such cases.[26]
Meanwhile there are a number of cases of mental blindness, especially for
written language, coupled with hemianopsia, usually of the rightward field
of view. These are all explicable by the breaking down, through disease,
of the connecting tracts between the occipital lobes and other parts
of the brain, especially those which go to the centres for speech in the
frontal and temporal regions of the left hemisphere. They are to be classed
among disturbances of conduction or of association; and nowhere
can I find any fact which should force us to believe that optical images
need[27] be lost in mental blindness, or that the cerebral
centres for such images are locally distinct from those for direct sensations
from the eyes.[28]
Where an object fails to be recognized by sight, it often happens that
the patient will recognize and name it as soon as he touches it with his
hand. This shows in an interes-[p.52] ting way how numerous the associative
paths are which all end by running out of the brain through the channel
of speech. The hand-path is open, though the eye-path be closed. When mental
blindness is most complete, neither sight, touch, nor sound avails to steer
the patient, and a sort of dementia which has been called asymbolia
or apraxia is the result. The commonest articles are not understood.
The patient will put his breeches on one shoulder and his hat upon the
other, will bite into the soap and lay his shoes on the table, or take
his food into his hand and throw it down again, not knowing what to do
with it, etc. Such disorder can only come from extensive brain-injury.[29]
The method of degeneration corroborates the other evidence localizing
the tracts of vision. In young animals one gets secondary degeneration
of the occipital regions from destroying an eyeball, and, vice versa,
degeneration of the optic nerves from destroying the occipital regions.
The corpora geniculata, thalami, and subcortical fibres leading to the
occipital lobes are also found atrophied in these cases. The phenomena
are not uniform, but are indisputable;[30] so that,
taking all lines of evidence together, the special connection of vision
with the occipital lobes is perfectly made out. It should be added that
the occipital lobes have frequently been found shrunken in cases of inveterate
blindness in man.
Hearing.
Hearing is hardly as definitely localized as sight. In the dog,
Luciani's diagram will show the regions which directly or indirectly affect
it for the worse when injured. As with sight, one-sided lesions produce
symptoms on both sides. The mixture of black dots and gray dots
in the diagram is meant to represent this mixture of 'crossed' and 'uncrossed'
connections, though of course no topographical exactitude is aimed at.
Of all the region, the temporal lobe is the most important part; yet permanent
absolute deafness did not [p.53] result in a dog of Luciani's, even from
bilateral destruction of both temporal lobes in their entirety.[31]
In the monkey, Ferrier and Yeo once found permanent deafness
to follow destruction of the upper temporal convolution (the one just below
the fissure of Sylvius in Fig.6) on both sides. Brown and Schaefer found,
on the contrary, that in several monkeys this operation failed to noticeably
affect the hearing. In one animal, indeed, both entire temporal lobes were
destroyed. After a week or two of depression of the mental faculties this
beast recovered and became one of the brightest monkeys possible, domineering
over all his mates, and admitted by all who saw him to have all his senses,
including hearing, 'perfectly acute.'[32] Terrible recriminations
have, as usual, ensued between the investigators, Ferrier denying that
Brown and Schaefer's ablations were complete,[33] Schaefer
that Ferrier's monkey was really deaf.[34] In this unsatisfactory
condition the subject must be left, although there seems no reason to doubt
that Brown and Schaefer's observation is the more important of the two.
In man the temporal lobe is unquestionably, the seat of the hearing
function, and the superior convolution adjacent to the sylvian fissure
is its most important part. The phenomena of aphasia show this. We studied
motor aphasia a few pages back; we must now consider sensory aphasia.[p.54]
Our knowledge of this disease has had three stages: we may talk
of the period of Broca, the period of Wernicke, and the period of Charcot.
What Broca's discovery was we have seen. Wernicke was the first to discriminate
those cases in which the patient can not even understand speech
from those in which he can understand, only not talk; and to ascribe the
former condition to lesion of the temporal lobe.[35]
The condition in question is word-deafness, and the disease is auditory
aphasia. The latest statistical survey of the subject is that by Dr.
Allen Starr.[36] In the seven cases of pure word-deafness
which he has collected, cases in which the patient could read, talk, and
write, but not understand what was said to him, the lesion was limited
to the first and second temporal convolutions in their posterior two thirds.
The lesion (in right-handed, i.e. left-brained, persons) is always on the
left side, like the lesion in motor aphasia. Crude hearing would not be
abolished, even were the left centre for it utterly destroyed ; the right
centre would still provide for that. But the linguistic use of hearing
appears bound up with the integrity of the left centre more or less exclusively.
Here it must be that words heard enter into association with the things
which they represent, on the one hand, and with the movements necessary
for pronouncing them, on the other. In a large majority of Dr. Starr's
fifty cases, the power either to name objects or to talk coherently was
impaired. This shows that in most of us (as Wernicke said) speech must
go on from auditory cues; that is, it must be that our ideas do not innervate
our motor centres directly, but only after first arousing the mental sound
of the words. This is the immediate stimulus to articulation; and
where the possibility of this is abolished by the destruction of its usual
channel in the left temporal lobe, the articulation must suffer. In the
few cases in which the channel is abolished with no bad effect on
speech we must suppose an idiosyncrasy. The patient must innervate his
speech-organs either from the corresponding portion of the other
hemisphere or directly from the centres of ideation, [p.55] those, namely,
of vision, touch, etc., without leaning on the auditory region. It is the
minuter analysis of the facts in the light of such individual differences
as these which constitutes Charcot's contribution towards clearing up the
subject.
Every namable thing, act, or relation has numerous properties, qualities,
or aspects. In our minds the properties of each thing, together with its
name, form an associated group. If different parts of the brain are severally
concerned with the several properties, and a farther part with the hearing,
and still another with the uttering, of the name, there must inevitably
be brought about (through the law of association which we shall later study)
such a dynamic connection amongst all these brain-parts that the activity
of anyone of them will be likely to awaken the activity of all the rest.
When we are talking as we think, the ultimate process is that of
utterance. If the brain-part for that be injured, speech is impossible
or disorderly, even though all the other brain-parts be intact: and this
is just the condition of things which, on page 37, we found to be brought
about by limited lesion of the left inferior frontal convolution. But back
of that last act various orders of succession are possible in the associations
of a talking man's ideas. The more usual order seems to be from the tactile,
visual, or other properties of the things thought-about to the sound of
their names, and then to the latter's utterance. But if in a certain individual
the thought of the look of an object or of the look of its
printed name be the process which habitually precedes articulation, then
the loss of the hearing centre will pro tanto not affect
that individual's speech. He will be mentally deaf, i.e. his understanding
of speech will suffer, but he will not be aphasic. In this way it is possible
to explain the seven cases of pure word-deafness which figure in
Dr. Starr's table.
If this order of association be ingrained and habitual in that individual,
injury to his visual centres will make him not only word-blind,
but aphasic as well. His speech will become confused in consequence of
an occipital lesion. Naunyn, consequently, plotting out on a diagram of
the hemisphere the 71 irreproachably reported cases of [p.56] aphasia which
he was able to collect, finds that the lesions concentrate themselves in
three places: first, on Broca's, centre; second, on Wernicke's ; third,
on the supra-marginal and angular gyri under which those fibres pass which
connect the visual centres with the rest of the brain [37](see
Fig. 17). With this result Dr. Starr's analysis of purely sensory cases
agrees.
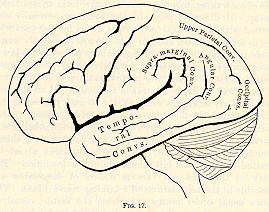
In a later chapter we shall again return to these differences in the
effectiveness of the sensory spheres in different individuals. Meanwhile
few things show more beautifully than the history of our knowledge of aphasia
how the sagacity and patience of many banded workers are in time certain
to analyze the darkest confusion into an orderly display.[38]
There is no 'centre of Speech' in the brain any more than there is a faculty
of Speech in the mind. The entire brain, more or less, is at work in a
man who uses language. The subjoined diagram, from Ross, shows the four
parts most critically concerned, and, in the light of our text, needs no
farther explanation (see Fig. 18).[p.57]
Smell.
Everything conspires to point to the median descending part of the temporal
lobes as being the organs of smell. Even Ferrier and Munk agree on the
hippocampal gyrus, though Ferrier restricts olfaction, as Munk does not
to the lobule or uncinate process of the convolution, reserving the rest
of it for touch.
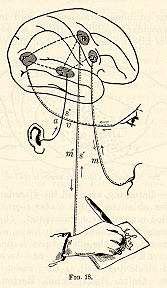
Anatomy and pathology also point to the hippocampal gyrus; but as the
matter is less interesting from the point of view of human psychology than
were sight and hearing, I will say no more, but simply add Luciani and
Seppili's diagram of the dog's smell-centre.[39]
Taste
Of [p.58] we know little that is definite.[sic] What little there is points
to the lower temporal regions again. Consult Ferrier as below.
Touch.
Interesting problems arise with regard to the seat of tactile and muscular
sensibility. Hitzig, whose experiments on dogs' brains fifteen years
ago opened the entire subject which we are discussing, ascribed the disorders
of motility observed after ablations of the motor region to a loss of what
he called muscular consciousness.
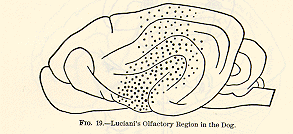
The animals do not notice eccentric positions of their limbs, will stand
with their legs crossed, with the affected paw resting on its back or hanging
over a table's edge, etc.; and do not resist our bending and stretching
of it as they resist with the unaffected paw. Goltz, Munk, Schiff, Herzen,
and others promptly ascertained an equal defect of cutaneous sensibility
to pain, touch, and cold. The paw is not withdrawn when pinched, remains
standing in cold water, etc. Ferrier meanwhile denied that there was any
true anaesthesia produced by ablations in the motor zone, and explains
the appearance of it as an effect of the sluggish motor responses of the
affected side.[40] Munk [41]and Schiff
[42], on the [p.59] contrary, conceive of the 'motor
zone' as essentially sensory, and in different ways explain the motor disorders
as secondary results of the anaesthesia which is always there. Munk calls
the motor zone the Fühlsphäre of the animal's limbs, etc., and
makes it coördinate with the Sehsphäre, the Hörsphäre,
etc., the entire cortex being, according to him, nothing but a projection-surface
for sensations, with no exclusively or essentially motor part. Such a view
would be important if true, through its bearings on the psychology of volition.
What is the truth? As regards the fact of cutaneous anaesthesia from motor-zone
ablations, all other observers are against Ferrier, so that he is probably
wrong in denying it. On the other hand, Munk and Schiff are wrong in making
the motor symptoms depend on the anaesthesia, for in certain rare
cases they have been observed to exist not only without insensibility,
but with actual hyperaesthesia of the parts.[43] The
motor and sensory symptoms seem, therefore, to be independent variables.
In monkeys the latest experiments are those of Horsley and Schaefer,[44]
whose results Ferrier accepts. They find that excision of the hippocampal
convolution produces transient insensibility of the opposite side of the
body, and that permanent insensibility is produced by destruction of its
continuation upwards above the corpus callosum, the so-called gyrus
fornicatus (the part just below the 'calloso-marginal fissure' in Fig.7).
The insensibility is at its maximum when the entire tract comprising both
convolutions is destroyed. Ferrier says that the sensibility of monkeys
is 'entirely unaffected' by ablations of the motor zone,[45]
and Horsley and Schaefer consider it by no means necessarily [p.60] abolished.[46]
Luciani found it diminished in his three experiments on apes.[47]
In man we have the fact that one-sided paralysis from disease of
the opposite motor zone may or may not be accompanied with anaesthesia
of the parts.
Luciani, who believes that the motor zone is also sensory, tries to
minimize the value of this evidence by pointing to the insufficiency with
which patients are examined. He himself believes that in dogs the tactile
sphere extends backwards and forwards of the directly excitable region,
into the frontal and parietal lobes (see Fig. 20). Nothnagel considers
that pathological evidence points in the same direction;[48]
and Dr. Mills, carefully reviewing the evidence, adds the gyri fornicatus
and hippocampi to the cutaneo-muscular region in man.[49]
If one compare Luciani's diagrams together (Figs. 14,16, 19, 20) one will
see that the entire parietal region of the dog's skull is common to the
four senses of sight, hearing, smell, and touch, including muscular feeling.
The corresponding region in the human brain (upper parietal and supra-marginal
gyri-see Fig. 17, p.56) seems to be a somewhat similar place of conflux.
Optical aphasias and motor and tactile disturbances all result from its
injury, especially when that is on the left side.[50]
The lower we go in the animal scale the [p.61] less differentiated the
functions of the several brain-parts seem to be.[51]
It may be that the region in question still represents in ourselves something
like this primitive condition, and that the surrounding parts, in adapting
themselves more and more to specialized and narrow functions, have left
it as a sort of carrefour through which they send currents and converse.
That it should be connected with musculo-cutaneous feeling is, however,
no reason why the motor zone proper should not be so connected too. And
the cases of paralysis from the motor zone with no accompanying anaesthesia
may be explicable without denying all sensory function to that region.
For, as my colleague Dr.James Putnam informs me, sensibility is always
harder to kill than motility, even where we know for a certainty that the
lesion affects tracts that are both sensory and motor. Persons whose hand
is paralyzed in its movements from compression of arm-nerves during sleep,
still feel with their fingers; and they may still feel in their feet when
their legs are paralyzed by bruising of the spinal cord. In a similar way,
the motor cortex might be sensitive as well as motor, and yet by this greater
subtlety (or whatever the peculiarity may be) in the sensory currents,
the sensibility might survive an amount of injury there by which the motility
was destroyed. Nothnagel considers that there are grounds for supposing
the muscular sense to be exclusively connected with the parietal
lobe and not with the motor zone. "Disease of this lobe gives pure ataxy
without palsy, and of the motor zone pure palsy without loss of muscular
sense.[52]" He fails, however, to convince more competent
critics than the present writer,[53] so I conclude with
them that as yet we have no decisive grounds for locating muscular and
cutaneous feeling apart. Much still remains to be learned about the relations
between musculo-cutaneous sensibility and the cortex, but one thing is
certain: that neither the occipital, the forward frontal, nor the temporal
lobes seem to have anything essential to do with it in man.[p.62] It is
knit up with the performances of the motor zone and of the convolutions
backwards and midwards of them. The reader must remember this
conclusion when we come to the chapter on the Will.
I must add a word about the connection of aphasia with the tactile sense.
On p.40 I spoke of those cases in which the patient can write but not read
his own writing. He cannot read by his eyes ; but he can read by the feeling
in his fingers, if he retrace the letters in the air. It is convenient
for such a patient to have a pen in hand whilst reading in this way, in
order to make the usual feeling of writing more complete.[54]
In such a case we must suppose that the path between the optical and the
graphic centres remains open, whilst that between the optical and the auditory
and articulatory centres is closed. Only thus can we understand how the
look of the writing should fail to suggest the sound of the words to the
patient's mind, whilst it still suggests the proper movements of graphic
imitation. These movements in their turn must of course be felt, and the
feeling of them must be associated with the centres for hearing and pronouncing
the words. The injury in cases like this where very special combinations
fail, whilst others go on as usual, must always be supposed to be of the
nature of increased resistance to the passage of certain currents of association.
If any of the elements of mental function were destroyed the incapacity
would necessarily be much more formidable. A patient who can both read
and write with his fingers most likely uses an identical 'graphic' centre,
at once sensory and motor, for both operations.
I have now given, as far as the nature of this book will allow, a complete
account of the present state of the localization-question. In its main
outlines it stands firm, though much has still to be discovered. The anterior
frontal lobes, for example, so far as is yet known, have no definite functions.
Goltz finds that dogs bereft of them both are incessantly in motion, and
excitable by every small stimulus. They are [p.63] irascible and amative
in an extraordinary degree, and their sides grow bare with perpetual reflex
scratching; but they show no local troubles of either motion or
sensibility. In monkeys not even this lack of inhibitory ability is shown,
and neither stimulation nor excision of the prefrontal lobes produces any
symptoms whatever. One monkey of Horsley and Schaefer's was as tame, and
did certain tricks as well, after as before the operation.[55]
It is probable that we have about reached the limits of what can be learned
about brain-functions from vivisecting inferior animals, and that we must
hereafter look more exclusively to human pathology for light. The existence
of separate speech and writing centres in the left hemisphere in man; the
fact that palsy from cortical injury is so much more complete and enduring
in man and the monkey than in dogs; and the farther fact that it seems
more difficult to get complete sensorial blindness from cortical ablations
in the lower animals than in man, all show that functions get more specially
localized as evolution goes on. In birds localization seems hardly to exist,
and in rodents it is much less conspicuous than in carnivora. Even for
man, however, Munk's way of mapping out the cortex into absolute areas
within which only one movement or sensation is represented is surely false.
The truth seems to be rather that, although there is a correspondence of
certain regions of the brain to certain regions of the body, yet the several
parts within each bodily region are represented throughout the whole
of the corresponding brain-region like pepper and salt sprinkled from the
same caster. This, however, does not prevent each 'part' from having its
focus at one spot within the brain-region. The various brain-regions
merge into each other in the same mixed way. As Mr.Horsley says: "There
are border centres, and the area of representation of the face merges into
that for the representation of the upper limb. If there was a focal lesion
at that point, you would have the movements of these two parts starting
together."[56] [p.64] The accompanying figure from Paneth
shows just how the matter stands in the dog.[57]
 I am speaking now
of localizations breadthwise over the brain-surface. It is conceivable
that there might be also localizations depthwise through the cortex. The
more superficial cells are smaller, the deepest layer of them is large;
and it has been suggested that the superficial cells are sensorial, the
deeper ones motor;[58] or that the superficial ones
in the motor region are correlated with the extremities of the organs to
be moved(fingers, etc.), the deeper ones with the more central segments
(wrist, elbow, etc.).[59] It need hardly be said that
all such theories are as yet but guesses.
I am speaking now
of localizations breadthwise over the brain-surface. It is conceivable
that there might be also localizations depthwise through the cortex. The
more superficial cells are smaller, the deepest layer of them is large;
and it has been suggested that the superficial cells are sensorial, the
deeper ones motor;[58] or that the superficial ones
in the motor region are correlated with the extremities of the organs to
be moved(fingers, etc.), the deeper ones with the more central segments
(wrist, elbow, etc.).[59] It need hardly be said that
all such theories are as yet but guesses.
We thus see that the postulate of Meynert and Jackson which we started
with on p.30 is on the whole most satisfactorily corroborated by subsequent
objective research. The highest centres do probably contain nothing
but arrangements for representing impressions and movements, and
other arrangements for coupling the activity of these arrangements
together.[60] Currents pouring in from the sense-organs
first excite some arrangements, [p.65] which in turn excite others, until
at last a motor discharge downwards of some sort occurs.
When this is once clearly grasped there remains little ground for keeping
up that old controversy about the motor zone, as to whether it is in reality
motor or sensitive. The whole cortex, inasmuch as currents run through
it, is both. All the currents probably have feelings going with them, and
sooner or later bring movements about. In one aspect, then, every centre
is afferent, in another efferent, even the motor cells of the spinal cord
having these two aspects inseparably conjoined. Marique,[61]
and Exner and Paneth[62] have shown that by cutting
round a 'motor' centre and so separating it from the influence of
the rest of the cortex, the same disorders are produced as by cutting it
out, so that really it is only the mouth of the funnel, as it were, through
which the stream of innervation, starting from elsewhere, pours;[63]
consciousness accompanying the stream, and being mainly of things seen
if the stream is strongest occipitally, of things heard if it is strongest
temporally, of things felt, etc., if the stream occupies most intensely
the 'motor zone.' It seems to me that some broad and vague formulation
like this is as much as we can safely venture on in the present state of
science; and in subsequent chapters I expect to give confirmatory reasons
for my view.
MAN'S CONSCIOUSNESS LIMITED TO THE HEMISPHERES
But is the consciousness which accompanies the activity of the cortex
the only consciousness that man has? or are his lower centres conscious
as well?
This is a difficult question to decide, how difficult one only learns
when one discovers that the cortex-consciousness itself of certain objects
can be seemingly annihilated in any good hypnotic subject by a bare wave
of his opera-[p.66] tor's hand, and yet be proved by circumstantial evidence
to exist all the while in a split-off condition, quite as 'ejective'[64]
to the rest of the subject's mind as that mind is to the mind of the bystanders.[65]
The lower centres themselves may conceivably all the while have a split-off
consciousness of their own, similarly ejective to the cortex-consciousness;
but whether they have it or not can never be known from merely introspective
evidence. Meanwhile the fact that occipital destruction in man may cause
a blindness which is apparently absolute (no feeling remaining either of
light or dark over one half of the field of view), would lead us to suppose
that if our lower optical centres, the corpora quadrigemina, and thalami,
do have any consciousness, it is at all events a consciousness which does
not mix with that which accompanies the cortical activities, and which
has nothing to do with our personal Self. In lower animals this may not
be so much the case. The traces of sight found (supra, p. 46) in dogs and
monkeys whose occipital lobes were entirely destroyed, may possibly have
been due to the fact that the lower centres of these animals saw, and that
what they saw was not ejective but objective to the remaining cortex, i.e.
it formed part of one and the same inner world with the things which that
cortex perceived. It may be, however, that the phenomena were due to the
fact that in these animals the cortical 'centres' for vision reach outside
of the occipital zone, and that destruction of the latter fails to remove
them as completely as in man. This, as we know, is the opinion of the experimenters
themselves. For practical purposes, nevertheless, and limiting the meaning
of the word consciousness to the personal self of the individual, we can
pretty confidently answer the question prefixed to this paragraph by saying
that the cortex is the sole organ of consciousness in man.[66]
If there [p.67] be any consciousness pertaining to the lower centres, it
is a consciousness of which the self knows nothing.
THE RESTITUTION OF FUNCTION.
Another problem, not so metaphysical, remains. The most general and
striking fact connected with cortical injury is that of the restoration
of function. Functions lost at first are after a few days or weeks
restored. How are we to understand this restitution ?
Two theories are in the field:
1) Restitution is due to the vicarious action either of the rest of
the cortex or of centres lower down, acquiring functions which until then
they had not performed;
2) It is due to the remaining centres (whether cortical or 'lower')
resuming functions which they had always had, but of which the wound had
temporarily inhibited the exercise. This is the view of which Goltz and
Brown-Séquard are the most distinguished defenders.
Inhibition is a vera causa, of that there can be no doubt. The
pneumogastric nerve inhibits the heart, the splanchnic inhibits the intestinal
movements, and the superior laryngeal those of inspiration. The nerve-irritations
which may inhibit the contraction of arterioles are innumerable, and reflex
actions are often repressed by the simultaneous excitement of other sensory
nerves. For all such facts the reader must consult the treatises on physiology.
What concerns us here is the inhibition exerted by different parts of the
nerve-centres, when irritated, on the activity of distant parts. The flaccidity
of a frog from 'shock,' for a minute or so after his medulla oblongata
is cut, is an inhibition from the seat of injury which quickly passes away.
What is known as 'surgical shock' (unconsciousness, pallor, dilatation
of splanchnic blood-vessels, and general syncope and collapse) in the human
subject is an inhibition which lasts a longer time. Goltz, Freusberg, and
others, cutting the spinal cord in dogs, proved that there were functions
inhibited still longer by the wound, but which reestablished themselves
ultimately if the animal was kept alive. The lumbar region of the cord
was thus found to contain independent vaso-motor centres, centres for erec-[p.68]
tion, for control of the sphincters, etc., which could be excited to activity
by tactile stimuli and as readily reinhibited by others simultaneously
applied.[67] We may therefore plausibly suppose that
the rapid reappearance of motility, vision, etc., after their first disappearance
in consequence of a cortical mutilation, is due to the passing off of inhibitions
exerted by the irritated surface of the wound. The only question is whether
all restorations of function must be explained in this one simple
way, or whether some part of them may not be owing to the formation of
entirely new paths in the remaining centres, by which they become 'educated'
to duties which they did not originally possess. In favor of an indefinite
extension of the inhibition theory facts may be cited such as the following:
In dogs whose disturbances due to cortical lesion have disappeared, they
may in consequence of some inner or outer accident reappear in all their
intensity for 24 hours or so and then disappear again.[68]
In a dog made half blind by an operation, and then shut up in the dark,
vision comes back just as quickly as in other similar dogs whose sight
is exercised systematically every day.[69] A dog which
has learned to beg before the operation recommences this practice quite
spontaneously a week after a double-sided ablation of the motor
zone.[70] Occasionally, in a pigeon (or even, it is
said, in a dog) we see the disturbances less marked immediately after the
operation than they are half an hour later.[71] This
would be impossible were they due to the subtraction of the organs which
normally carried them on. Moreover the entire drift of recent physiological
and pathological speculation is towards enthroning inhibition as an ever-present
and indispensable condition of orderly activity. We shall see how great
is its importance, in the chapter on the Will. Mr. Charles Mercier considers
that no muscular contraction, once begun, would ever stop without it, short
of exhaustion [p.69] of the system;[72] and Brown-Séquard
has for years been accumulating examples to show how far its influence
extends.[73] Under these circumstances it seems as if
error might more probably lie in cutailing its sphere too much than in
stretching it too far as an explanation of the phenomena following cortical
lesion.[74]
On the other hand, if we admit no re-education of centres, we
not only fly in the face of an a priori probability, but we find
ourselves compelled by facts to suppose an almost incredible number of
functions natively lodged in the centres below the thalami or even
in those below the corpora quadrigemina. I will consider the a
priori objection after first taking a look at the facts which I have
in mind. They confront us the moment we ask ourselves just which
are the parts which perform the functions abolished by an operation
after sufficient time has elapsed for restoration to occur?.
The first observers thought that they must be the corresponding parts
of the opposite or intact hemisphere. But as long ago as 1875 Carville
and Duret tested this by cutting out the fore-leg-centre on one side, in
a dog, and then, after waiting till restitution had occurred, cutting it
out on the opposite side as well. Goltz and others have done the same thing.[75]
If the opposite side were really the seat of the restored function, the
original palsy should have appeared again and been permanent. But it did
not appear at all; there appeared only a palsy of the hitherto unaffected
side. The next supposition is that the parts surrounding the cut-out
region learn vicariously to perform its duties. But here, again, experiment
seems to upset the hypothesis, so far as the motor zone goes at least;
for we may wait till motility has returned in the affected limb, and then
both irritate the [p.70] cortex surrounding the wound without exciting
the limb to movement, and ablate it, without bringing back the vanished
palsy.[76] It would accordingly seem that the cerebral
centres below the cortex must be the seat of the regained activities.
But Goltz destroyed a dog's entire left hemisphere, together with the corpus
striatum and the thalamus on that side, and kept him
alive until a surprisingly small amount of motor and tactile disturbance
remained.[77] These centres cannot here have accounted
for the restitution. He has even, as it would appear,[78]
ablated both the hemispheres of a dog, and kept him alive 51 days, able
to walk and stand. The corpora striata and thalami in this dog were also
practically gone. In view of such results we seem driven, with M.Francois-Franck,[79]
to fall back on the ganglia lower still, or even on the spinal
cord as the 'vicarious' organ of which we are in quest. If the abeyance
of function between the operation and the restoration was due exclusively
to inhiibition, then we must suppose these lowest centres to be in reality
extremely accomplished organs. They must always have done what we now find
them doing after function is restored, even when the hemispheres were intact.
Of course this is conceivably the case; yet it does not seem very plausible.
And the a priori considerations which a moment since I said I should
urge, make it less plausible still.
For, in the first place, the brain is essentially a place of currents,
which run in organized paths. Loss of function can only mean one of two
things, either that a current can no longer run in, or that if it runs
in, it can no longer run out, by its old path. Either of these inabilities
may come from a local ablation; and 'restitution' can then only mean that,
in spite of a temporary block, an inrunning current has at last become
enabled to flow out by its old path again-e.g., the sound of 'give your
paw' discharges after some [p.71] weeks into the same canine muscles into
which it used to discharge before the operation. As far as the cortex itself
goes, since one of the purposes for which it actually exists is the
production of new paths,[80] the only question before
us is: Is the formation of these particular 'vicarious' paths too
much to expect of its plastic powers? It would certainly be too much to
expect that a hemisphere should receive currents from optic fibres whose
arriving-place within it is destroyed, or that it should discharge
into fibres of the pyramidal strand if their place of exit is broken
down. Such lesions as these must be irreparable within that hemisphere.
Yet even then, through the other hemisphere, the corpus callosum,
and the bilateral connections in the spinal cord, one can imagine some
road by which the old muscles might eventually be innervated by the same
incoming currents which innervated them before the block. And for all minor
interruptions, not involving the arriving-place of the 'cortico-petal'
or the place of exit of the 'cortico-fugal' fibres, roundabout paths of
some sort through the affected hemisphere itself must exist, for every
point of it is, remotely at least, in potential communication with every
other point. The normal paths are only paths of least resistance. If they
get blocked or cut, paths formerly more resistant become the least resistant
paths under the changed conditions. It must never be forgotten that a current
that runs in has got to run out somewhere; and if it only once succeeds
by accident in striking into its old place of exit again, the thrill of
satisfaction which the consciousness connected with the whole residual
brain then receives will reinforce and fix the paths of that moment and
make them more likely to be struck into again. The resultant feeling that
the old habitual act is at last successfully back again, becomes itself
a new stimulus which stamps all the existing currents in. It is matter
of experience that such feelings of successful achievement do tend to fix
in our memory whatever processes have led to them; and we shall have [p.72]
a good deal more to say upon the subject when we come to the Chapter on
the Will.
My conclusion then is this: that some of the restitution of function
(especially where the cortical lesion is not too great) is probably due
to genuinely vicarious function on the part of the centres that remain;
whilst some of it is due to the passing off of inhibitions. In other words,
both the vicarious theory and the inhibition theory are true in their measure.
But as for determining that measure, or saying which centres are vicarious,
and to what extent they can learn new tricks, that is impossible at present.
FINAL CORRECTION OF THE MEYNERT SCHEME.
And now, after learning all these facts, what are we to think of the
child and the candle-flame, and of that scheme which provisionally imposed
itself on our acceptance after surveying the actions of the frog? (Cf.
pp. 25-6, supra.) It will be remembered that we then considered
the lower centres en masse as machines for responding to present
sense-impressions exclusively, and the hemispheres as equally exclusive
organs of action from inward considerations or ideas; and that, following
Meynert, we supposed the hemispheres to have no native tendencies to determinate
activity, but to be merely superadded organs for breaking up the various
reflexes performed by the lower centres, and combining their motor and
sensory elements in novel ways. It will also be remembered that I prophesied
that we should be obliged to soften down the sharpness of this distinction
after we had completed our survey of the farther facts. The time has now
come for that correction to be made.
Wider and completer observations show us both that the lower centres
are more spontaneous, and that the hemispheres are more automatic, than
the Meynert scheme allows. Schrader's observations in Goltz's Laboratory
on hemisphereless frogs[81] and pigeons[82]
give an idea quite different from the picture of these creatures which
is classically current. Steiner's[83] observations on
frogs [p.73] already went a good way in the same direction, showing, for
example, that locomotion is a well-developed function of the medulla oblongata.
But Schrader, by great care in the operation, and by keeping the frogs
a long time alive, found that at least in some of them the spinal cord
would produce movements of locomotion when the frog was smartly roused
by a poke, and that swimming and croaking could sometimes be performed
when nothing above the medulla oblongata remained.[84]
Schrader's hemisphereless frogs moved spontaneously, ate flies, buried
themselves in the ground, and in short did many things which before his
observations were supposed to be impossible unless the hemispheres remained.
Steinert[85] and Vulpian have remarked an even greater
vivacity in fishes deprived of their hemispheres. Vulpian says of his brainless
carps[86] that three days after the operation one of
them darted at food and at a knot tied on the end of a string, holding
the latter so tight between his jaws that his head was drawn out of water.
Later, "they see morsels of white of egg; the moment these sink through
the water in front of them, they follow and seize them, sometimes after
they are on the bottom, sometimes before they have reached it. In capturing
and swallowing this food they execute just the same movements as the intact
carps which are in the same aquarium. The only difference is that they
seem to see them at less distance, seek them with less impetuosity and
less perseverance in all the points of the bottom of the aquarium, but
they struggle (so to speak) sometimes with the sound carps to grasp the
morsels. It is certain that they do not confound these bits of white of
egg with other white bodies, small pebbles for example, which are at the
bottom of the water. The same carp which, three days after operation, seized
the knot on a piece of string, no longer snaps at it now, but if one brings
it near her, she draws away from it by swimming backwards before it comes
into contact with [p.74] her mouth."[87] Already on
pp.9-10,as the reader may remember, we instanced those adaptations of conduct
to new conditions, on the part of the frog's spinal cord and thalami, which
led Pfüger and Lewes on the one hand and Goltz on the other to locate
in these organs an intelligence akin to that of which the hemispheres are
the seat.
When it comes to birds deprived of their hemispheres, the evidence that
some of their acts have conscious purpose behind them is quite as persuasive.
In pigeons Schrader found that the state of somnolence lasted only three
or four days, after which time the birds began indefatigably to walk about
the room. They climbed out of boxes in which they were put, jumped over
or flew up upon obstacles, and their sight was so perfect that neither
in walking nor flying did they ever strike any object in the room. They
had also definite ends or purposes, flying straight for more convenient
perching places when made uncomfortable by movements imparted to those
on which they stood; and of several possible perches they always chose
the most convenient. "If we give the dove the choice of a horizontal bar
(Reck) or an equally distant table to fly to, she always gives decided
preference to the table. Indeed she chooses the table even if it is several
meters farther off than the bar or the chair." Placed on the back of a
chair, she flies first to the seat and then to the floor, and in general
,"will forsake a high position, although it give her sufficiently firm
support, and in order to reach the ground will make use of the environing
objects as intermediate goals of flight, showing a perfectly correct judgment
of their distance. Although able to fly directly to the ground, she prefers
to make the journey in successive stages. . . . Once on the ground, she
hardly ever rises spontaneously into the air."[88]
Young rabbits deprived of their hemispheres will stand, run, start at
noises, avoid obstacles in their path, and give responsive cries
of suffering when hurt. Rats will do the same, and throw themselves moreover
into an attitude of defence. Dogs never survive such an operation if performed
at once. But Goltz's latest dog, mentioned on p.[p.75] 70, which is said
to have been kept alive for fifty-one days after both hemispheres had been
removed by a series of ablations and the corpora striata and thalami had
softened away, shows how much the mid-brain centres and the cord can do
even in the canine species. Taken together, the number of reactions shown
to exist in the lower centres by these observations make out a pretty good
case for the Meynert scheme, as applied to these lower animals. That scheme
demands hemispheres which shall be mere supplements or organs of repetition,
and in the light of these observations they obviously are so to a great
extent. But the Meynert scheme also demands that the reactions of the lower
centres shall all be native, and we are not absolutely sure that
some of those which we have been considering may not have been acquired
after the injury; and it furthermore demands that they should be machine-like,
whereas the expression of some of them makes us doubt whether they may
not be guided by an intelligence of low degree.
Even in the lower animals, then, there is reason to soften down that
opposition between the hemispheres and the lower centres which the scheme
demands. The hemispheres may, it is true, only supplement the lower centres,
but the latter resemble the former in nature and have some small amount
at least of 'spontaneity' and choice.
But when we come to monkeys and man the scheme well-nigh breaks down
altogether; for we find that the hemispheres do not simply repeat voluntarily
actions which the lower centres perform as machines. There are many functions
which the lower centres cannot by themselves perform at all. When the motor
cortex is injured in a man or a monkey genuine paralysis ensues, which
in man is incurable, and almost or quite equally so in the ape. Dr. Seguin
knew a man with hemi-blindness, from cortical injury, which had persisted
unaltered for twenty-three years. 'Traumatic inhibition' cannot possibly
account for this. The blindness must have been an 'Ausfallserscheinung,'
due to the loss of vision's essential organ. It would seem, then, that
in these higher creatures the lower centres must be less adequate than
they are farther down in the zoological scale; and that even for certain
elementary [p.76] combinations of movement and impression the co-operation
of the hemispheres is necessary from the start. Even in birds and dogs
the power of eating properly is lost when the frontal lobes are
cut off.[89]
The plain truth is that neither in man nor beast are the hemispheres
the virgin organs which our scheme called them. So far from being unorganized
at birth, they must have native tendencies to reaction of a determinate
sort.[90] These are the tendencies which we know as
emotions and instincts, and which we must study with some
detail in later chapters of this book. Both instincts and emotions are
reactions upon special sorts of objects of perception; they depend
on the hemispheres; and they are in the first instance reflex, that is,
they take place the first time the exciting object is met, are accompanied
by no forethought or deliberation, and are irresistible. But they are modifiable
to a certain extent by experience, and on later occasions of meeting the
exciting object, the instincts expecially have less of the blind impulsive
character which they had at first. All this will be explained at some length
in Chapter XXIV. Meanwhile we can say that the multiplicity of emotional
and instincitive reactions in man, together with his extensive associative
power, permit of extensive recouplings of the original sensory and motor
partners. The consequences of one instinctive reaction often prove
to be the inciters of an opposite reaction, and being suggested
on later occasions by the original object, may then suppress the first
reaction altogether, just as in the case of the child and the flame. For
this education the hemispheres do not need [p.77] to be tabuloe rasoe
at first, as the Meynert scheme would have them; and so far from their
being educated by the lower centres exclusively, they educate themselves.[91]
We have already noticed the absence of reactions from fear and hunger
in the ordinary brainless frog. Schrader gives a striking account of the
instinctless condition of his brainless pigeons, active as they were in
the way of locomotion and voice. "The hemisphereless animal moves in a
world of bodies which ... are all of equal value for him.... He is, to
use Goltz's apt expression, impersonal.... Every object is for him
only a space-occupying mass, he turns out of his path for an ordinary pigeon
no otherwise than for a stone. He may try to climb over both. All authors
agree that they never found any difference, whether it was an inanimate
body, a cat, a dog, or a bird of prey which came in their pigeon's way.
The creature knows neither friends nor enemies, in the thickest company
it lives like a hermit. The languishing cooing of the male awakens no more
impression than the rattling of the peas, or the call-whistle which in
the days before the injury used to make the birds hasten to be fed. Quite
as little as the earlier observers have I seen hemisphereless she-birds
answer the courting of the male. A hemisphereless male will coo all day
long and show distinct signs of sexual excitement, but his activity
is without any object, it is entirely indifferent to him whether the
she-bird be there or not. If one is placed near him, he leaves
her unnoticed.... As the male pays no attention to the female, so she pays
none to her young. The brood may follow the mother ceaselessly calling
for food, but they might as well ask it from a stone.... The hemi[p.78]
sphereless pigeon is in the highest degree tame, and fears man as
little as cat or bird of prey."[92]
Putting together now all the facts and reflections which we have been
through, it seems to me that we can no longer hold strictly to
the Meynert scheme. If anywhere, it will apply to the lowest animals;
but in them especially the lower centres seem to have a degree of spontaneity
and choice. On the whole, I think that we are driven to substitute for
it some such general conception as the following, which allows for zoological
differences as we know them, and is vague and elastic enough to receive
any number of future discoveries of detail.
CONCLUSION.
All the centres, in all animals, whilst they are in one aspect mechanisms,
probably are, or at least once were, organs of consciousness in another,
although the consciousness is doubtless much more developed in the hemispheres
than it is anywhere else. The consciousness must everywhere prefer
some of the sensations which it gets to others; and if it can remember
these in their absence, however dimly, they must be its ends of desire.
If, moreover, it can identify in memory any motor discharges which may
have led to such ends, and associate the latter with them, then
these motor discharges themselves may in turn become desired as means.
This is the development of will; and its realization must of course
be proportional to the possible complication of the consciousness. Even
the spinal cord may possibly have some little power of will in this sense,
and of effort towards modified behavior in consequence of new experiences
of sensibility.[93]
[p.79] All nervous centres have then in the first instance one essential
function, that of 'intelligent' action. They feel, prefer one thing to
another, and have 'ends.' Like all other organs, however, they evolve
from ancestor to descendant, and their evolution takes two directions the
lower centres passing downwards into more unhesitating automatism, and
the higher ones upwards into larger intellectuality.[94]
Thus it may happen that those functions which can safely grow uniform and
fatal become least accompanied by mind, and that their organ, the spinal
cord, becomes a more and more soulless machine; whilst on the contrary
those functions which it benefits the animal to have adapted to delicate
environing variations pass more and more to the hemispheres, whose anatomical
structure and attendant consciousness grow more and more elaborate as zoological
evolution proceeds. In this way it might come about that in man and the
monkeys the basal ganglia should do fewer things by themselves than they
can do in dogs, fewer in dogs than in rabbits, fewer in rabbits than in
hawks,[95] fewer in hawks than in pigeons, fewer in
pigeons than in frogs, fewer in frogs than in fishes, and that the hemispheres
should correspondingly do more. This passage of functions forward to the
ever-enlarging hemispheres would be itself one of the evolutive changes,
to be explained like the development of the hemispheres themselves, either
by fortunate variation or by inherited effects of use. The reflexes, on
this view, upon which the education of our human hemispheres depends, would
not be due to the basal ganglia [p.80] alone. They would be tendencies
in the hemispheres themselves, modifiable by education, unlike the reflexes
of the medulla oblongata, pons, optic lobes and spinal cord. Such cerebral
reflexes, if they exist, form a basis quite as good as that which the Meynert
scheme offers, for the acquisition of memories and associations which may
later result in all sorts of 'changes of partners' in the psychic world.
The diagram of the baby and the candle (see page 25) can be re-edited,
if need be, as an entirely cortical transaction. The original tendency
to touch will be a cortical instinct; the burn will leave an image in another
part of the cortex, which, being recalled by association, will inhibit
the touching tendency the next time the candle is perceived, and excite
the tendency to withdraw-so that the retinal picture will, upon that next
time, be coupled with the original motor partner of the pain. We thus get
whatever psychological truth the Meynert scheme possesses without entangling
ourselves on a dubious anatomy and physiology.
Some such shadowy view of the evolution of the centres, of the relation
of consciousness to them, and of the hemispheres to the, other lobes, is,
it seems to me, that in which it is safest to indulge. If it has no other
advantage, it at any rate makes us realize how enormous are the gaps in
our knowledge, the moment we try to cover the facts by any one formula
of a general kind.
Footnotes
[1] It should be said that this particular cut commonly
proves fatal. The text refers to the rare cases which survive.
[2] I confine myself to the frog for simplicity's sake.
In higher animals, especially the ape and man, it would seem as if not
only determinate combinations of muscles, but limited groups or even single
muscles could be innervated from the hemispheres.
[3] I hope that the reader will take no umbrage at
my so mixing the physical and mental, and talking of reflex acts and hemispheres
and reminiscences in the same breath, as if they were homogeneous quantities
and factors of one causal chain. I have done so deliberately; for although
I admit that from the radically physical point of view it is easy to conceive
of the chain of events amongst the cells and fibres as complete in itself,
and that whilst so conceiving it one need make no mention of ideas,' I
yet suspect that point of view of being an unreal abstraction. Reflexes
in centres may take place even where accompanying feelings or ideas guide
them. In another chapter I shall try to show reasons for not abandoning
this common-sense position; meanwhile language lends itself so much more
easily to the mixed way of describing , that I will continue to employ
the latter. The more radical-minded reader can alway read 'ideational process'
for idea'.
[4] I shall call it hereafter for shortness 'the Meynert
scheme;' for the child-and-flame example, as well as the whole general
notion that the hemispheres are a supernumerary surface for the projection
and association of sensations and movements natively coupled in the centres
below, is due to Th. Meynert, the Austrian anatomist. For a popular account
of his views, see his pamphlet 'Zur Mechanik des Gehirnbaues,' Vienna,
1874. His most recent development of them is embodied in his 'Psychiatry,'
a clinical treatise on diseases of the forebrain, translated by B.Sachs,
New York, 1885.
[5] Geschichte des Materialismus, 2d ed., II. p 345.
[6] West Riding Asylum Reports, 1876, p. 267.
[7] For a thorough discussion of the various objections,
see Ferrier's 'Functions of the Brain,' 2d ed., pp. 227-234, and Franois-Franck's
'Leons sur les Fonctions Motrices du Cerveau'(1887), Leon 31. The most
minutely accurate experiments on irritation of cortical points are those
of Paneth, in Pflüger's Archiv, vol 37, p. 528.-Recently the skull
has been fearlessly opened by surgeons, and operations upon the human brain
performed, sometimes with the happiest results. In some of these operations
the cortex has been electrically excited for the purpose of more exactly
localizing the spot, and the movements first observed in dogs and monkeys
have then been verified in men.
[8] J. Loeb: 'Beiträge zur Physiologie des Grosshirns;'
Pflüger's Arciv, XXXIX. 293. I simplify the author's statement.
[9] Goltz: Pflüger's Archiv, XLII. 419.
[10] 'Hemiplegia' means one-sided palsy.
[11] Philosophical Transactions, vol. 179, pp. 6,
10(1888). In a later paper (ibid. p. 205) Messrs. Beevor and Horsley
go into the localization still more minutely, showing spots from which
single muscles or single digits can be made to contract.
[12] Nothnagel und Naunyn : Die Localization in den
Gehirnkrankheiten (Wiesbaden, 1887), p.34
[13] An accessible account of the history of our knowledge
of motor aphasia is in W.A. Hammond's 'Treatise on the Diseases of the
Nervous System,' chapter VII.
[14] The history up to 1885 may be found in A.Christiani:
Zur Physiologie des Gehirnes (Berlin, 1885)
[15] Pflüger's Archiv, vol.44, p.176. Munk (Berlin
Academy Sitzsungsberichte, 1889, XXXI) returns to the charge, denying the
extirpations of Schrader to be complete: "Microscopic portions of the Sehsphäre
must remain."
[16] A.Christiani: Zur Physiol. D. Gehirnes (Berlin,
1885), chaps. II, III, IV. H. Munk: Berlin Akad. Stzgsb. 1884, XXIV.
[17] Luciani und Seppili: Die Functions-Localization
auf der Grosshirnrinde (Deutsch von Fraenkel), Leipzig, 1886, Dogs M, N,
and S. Goltz in Pflüger's Archiv, vol.34, pp. 490-6; vol. 42, p. 454.
Cf. also Munk: Berlin Akad. Stzgsb. 1886, VII, VIII, pp. 113-121, and Loeb:
Pflüger's Archiv, vol. 39, p. 337.
[18] Berlin Akad. Sitzungsberichte, 1886, VII, VIII,
p. 124.
[19] H. Munk: Functionen der Grosshirnrinde (Berlin,
1881), pp. 36-40. Ferrier: Functions, etc., 2d ed., chap. IX, pt. I. Brown
and Schaefer: Philos. Transactions, vol. 179, p. 321. Luciani u. Seppili,
op. Cit. Pp. 131-138. Lannegrace found traces of sight with both occipital
lobes destroyed, and in one monkey even when angular gyri and occipital
lobes were destroyed altogether. His paper is in the Archives de Médecine
Expérimentale for January and March, 1889. I only know it from the
abstract in the Neurologisches Centralblatt, 1889, pp. 108-420. The reporter
doubts the evidence of vision in the monkey. It appears to have consisted
in avoiding obstacles and in emotional disturbance in the presence of men.
[20] Localization of Cerebral Disease (1878), pp.
117-8.
[21] For cases see Flechsig : Die Leitungsbahnen in
Gehirn u. Rückenmark (Leipzig, 1876), pp. 112, 272; Exner's Untersuchungen,
etc., p. 83; Ferrier's Localization, etc., p. 11; Francois-Franck's Cerveau
Moteur, p. 63, note.
[22] E. C. Seguin: Hemianopsia of Cerebral Origin,
in Journal of Nervous and Mental Disease, vol. XIII. P. 30. Nothnagel und
Naunyn: Ueber die Localization der Gehirnkrankheiten (Wiesbaden, 1887),
p. 10.
[23] Die Seelenblindheit, etc., p. 51 ff. The mental
blindness was in this woman's case moderate in degree.
[24] Archiv f. Psychiatrie, vol. 21, p. 222.
[25] Nothnagel (loc. cit. p.22) says: "Dies trifft
aber nicht zu." He gives, however, no case in support of his opinion
that double-sided cortical lesion may make one stone-blind and yet not
destroy one's visual images; so that I do not know whether it is an observation
of fact or an a priori assumption.
[26] In a case published by C.S. Freund: Archiv f.
Psychiatrie, vol. XX, the occipital lobes were injured, but their cortex
was not destroyed, on both sides. There was still vision. Cf. pp. 291-5.
[27] I say 'need,' for I do not of course deny the
possible coexistence of the two symptoms. Many a brain-lesion might
block optical associations and at the same time impair optical imagination,
without entirely stopping vision. Such a case seems to have been the remarkable
on from Charcot which I shall give rather fully in the chapter on Imagination.
[28] Freund (in the article cited above "Ueber optisched
Aphasie und Seelenblindheit') and Bruns ('Ein Fall von Alexie,' etc., in
the Neurologisches Centralblatt for 1888, pp. 581, 509) explain their cases
by brokendown conduction. Wilbrand, whose painstaking monograph on mental
blindness was referred to a moment ago, gives none but a priori
reasons for his belief that the optical 'Erinnerungsfeld' must be locally
distinct from the Wahrnehmungsfeld (cf. pp. 84, 93). The a priori
reasons are really the other way. Mauthner ('Gehirn u. Auge' (1881), p.
487 ff.) tries to show that the 'mental blindness' of Munk's dogs and apes
after occipital mutilation was not such, but real dimness of sight. The
best case of mental blindness yet reported is that by Lissauer, as below.
The reader will also do well to read Bernard: De 1 Aphasie(1881) chap.
V; Ballet: Le Langage Intérieur (1886), chap. VIII; and Jas. Ross's
little book on Aphasia (1887), p. 74.
[29] For a case see Wernicke's Lehrb. D. Gehirnkrankheiten,
vol. II. p. 554(1881).
[30] The latest account of them is the paper Über
die optischen Centren u. Bahnen' by von Monakow in the Archiv für
Psychiatrie, vol. XX. p. 714.
[31] Die Functions-Localization, etc., Dog X; see
also p. 161.
[32] Philos. Trans., vol. 179, p. 312.
[33] Brain, vol. XI. p. 10.
[34] Ibid. p. 147.
[35] Der aphasische Symptomencomplex (1874). See in
Fig. 11 the convolution marked WERNICKE.
[36] 'The Pathology of Sensory Aphasia,' 'Brain,'
July, 1889.
[37] Nothnagel und Naunyn: op. cit., plates.
[38] Ballet's and Bernard's works cited on p. 51 are
the most accessible documents of Charcot's school. Bastian's book on the
Brain as an Organ of Mind(last three chapters) is also good.
[39] For details, see Ferrier's 'Functions,' chap,
IX. Pt. III, and Chas. K. Mills: Transactions of Congress of American Physicians
and Surgeons, 1888, vol. I. p. 278.
[40] Functions of the Brain, chap. X. 14.
[41] Uber die Functionen d. Grosshirnrinde (1881),
p. 50.
[42] Lezioni di Fisiologia sperimentale sul sistema
nervoso encefalico (1873), p. 527 ff. Also 'Brain,' vol. IX. p. 298.
[43] Bechterew (Pflüger's Archiv., vol. 35, p.
137) found no anaesthesia in a cat with motor symptoms from ablation
of sigmoid gyrus. Luciani got hyperaesthesia coexistent with cortical motor
defect in a dog, by simultaneously hemisecting the spinal cord (Luciani
u. Seppili, op. cit. p. 234). Goltz frequently found hyperaesthesia
of the whole body to accompany motor defect after ablation of both frontal
lobes, and he once found it after ablating the motor zone (Pflüger's
Archiv, vol. 34, p. 471).
[44] Philos. Transactions, vol. 179, p. 20 ff.
[45] Functions, p. 375.
[46] Pp. 15-17.
[47] Luciani u. Sepplili, op. cit. pp. 275-288.
[48] Op. cit. p. 18.
[49] Trans. Of Congress, etc., p. 272.
[50] See Exner's Unters. üb . Localization, plate
XXV.
[51] Cf. Ferrier's Functions, etc., chap. IV and chap.
X, 6 to 9.
[52] Op. cit. p.17.
[53] E.g. Starr, loc. cit. p. 272; Leyden,
Beiträge zur Lehre v. d. Localization im Gehirn(1888), p. 72.
[54] Bernard, op. cit. p. 84.
[55] Philos. Trans., vol. 179, p. 3.
[56] Trans. Of Congress of Am. Phys. And Surg. 1888,
vol. I.p. 343. Beevor and Horsley's paper on electric stimulation of the
monkey's brain is the most beautiful work yet done for precision. See Phil.
Trans., vol. 179, p. 205, especially the plates.
[57] Pflüger's Archiv, vol. 37, p. 523 (1885).
[58] By Luys in his generally preposterous book 'The
Brain'; also by Horsley.
[59] C. Mercier: The Nervous System and the Mind,
p. 124.
[60] The frontal lobes as yet remain a puzzle. Wundt
tries to explain them as an organ of 'apperception' (Grundzüge d.
Physiologischen Psychologie, 3d ed., vol. I. p. 233 ff.), but I confess
myself unable to apprehend clearly the Wundtian philosophy so far as this
word enters into it, so must be contented with this bare reference.- Until
quite recently it was common to talk of an 'ideational centre' as of something
distinct from the aggregate of other centres. Fortunately this custom is
already on the wane.
[61] Rech.Exp. sur le Fonctionnement des Centres Psycho-moteurs(Burssels,
1885).
[62] Pflüger's Archiv, vol. 44, p. 544.
[63] I ought to add, however, that Franois-Franck(Fonctions
Motrices, p. 370) got , in two dogs and a cat, a different result from
this sort of 'circumvallation.'
[64] For this word, see T.K. Clifford's Lectures and
Essays(1879), vol. II p. 72.
[65] See below, Chapter VIII.
[66] Cf. Ferrier's Functions, pp. 120, 147, 414. See
also Vulpian: Leons sur la Physiol. Du Syst. Nerveux, p. 548; Luciani u.
Seppili, op. cit. pp. 404-5; H. Maudsley: Physiology of Mind (1876),
pp. 138 ff., 197 ff., and 241 ff. In G.H. Lewes's Physical Basis of Mind,
Problem IV: 'The Reflex Theory,' a very full history of the question is
given.
[67] Goltz: Pflüger's Archiv, vol. 8, p. 460;
Freusberg: ibid. vol. 10, p. 174.
[68] Goltz: Verrichtungen des Grosshirns. p. 73.
[69] Loeb: Pflüger's Archiv, vol 39, p. 276.
[70] Ibid, p. 289.
[71] Schrader: ibid. vol. 44, p. 218.
[72] The Nervous System and the Mind (1888), chaps.
III, VI; also in Brain, vol. XI. p. 361.
[73] Brown-Séquard has given a résumé
of his opinions in the Archives de Physiologie for Oct. 1889, 5me, Série,
vol. I. p 751.
[74] Goltz first applied the inhibition thoery to
the brain in his 'Verrichtungen des Grosshirns,'p. 39 ff. On the general
philosophy of Inhibition the reader may consult Brunton's ' Pharmakology
and Therapeutics,' p. 154 ff., and also 'Nature,' vol. 27, p. 419 ff.
[75] E.g. Herzen, Herman u. Schwalbe's Jahres-bericht
for 1886, Physiol. Abth. P. 38. (Experiments on new-born puppies.)
[76] Franois-Franck: op.cit. p. 382. Results are somewhat
contradictory.
[77] Pflüger's Archiv, vol. 42, p. 419.
[78] Neurologisches Centralblatt, 1889, p. 372.
[79] Op. cit. p. 387. See pp. 378 to 388 for
a discussion of the whole question. Compare also Wundt's Physiol. Psych.,
3d ed., I. 225 ff., and Luciani u. Seppili, pp. 243, 293.
[80] The Chapters on Habit, Association, Memory, and
Perception will change our present preliminary conjecture that that is
one of its essential uses, into an unshakable conviction.
[81] Pflüger's Archiv, vol. 41, p. 75 (1887).
[82] Ibid., vol. 44, p. 175 (1889).
[83] Untersuchungen über die Physiologie des
Froschirns, 1885.
[84] Loc. cit. pp. 80, 82-3. Schrader also
found a biting-reflex developed when the medulla oblongata is cut
through just behind the cerebellum.
[85] Berlin Akad. Sitzungsberichte for 1886.
[86] Comptes Rendus, vol. 102, p. 90.
[87] Comptes Rendus de l'Acad. D. Sciences, vol. 102,
p. 1530
[88] Loc. cit. p. 216.
[89] Goltz: Pflüger's Archiv, vol. 42, p. 447;
Schrader : ibid. vol. 44, p. 219 ff. It is possible that this symptom
may be an effect of traumatic inhitition however.
[90] A few years ago one of the strongest arguements
for the theory that the hemispheres are purely supernumerary was Soltmann's
often-quoted observation that in new-born puppies the motor zone of the
cortex is not excitable by electricity and only becomes so in the course
of a fortnight, presumably after the experiences of the lower centres have
educated it to motor duties. Paneth's later observations, however, seem
to show that Soltmann may have been misled through overnarcotizing his
victims (Pflüger's Archiv, vol. 37, p. 202). In the Neurologisches
Centralblatt for 1889, p. 513, Bechterw returns to the subject on Soltmann's
side without however, noticing Paneth's work.
[91] Münsterberg (Die Willenshandlung, 1888,
p. 134) challenges Meynert's scheme in toto, saying that whilst
we have in our personal experience plenty of examples of acts which were
at first voluntary becoming secondarily automatic and reflex, we have no
conscious record of a single originally reflex act growing voluntary. -As
far as conscious record is concerned, we could not possibly have it even
if the Meynert scheme were wholly true, for the education of the hemispheres
which that scheme postulates must in the nature of things antedate recollection.
But it seems to me that Münsterberg's rejection of the scheme may
possibly be correct as regards reflexes from the lower centres.
Everywhere in this department of psychogenesis we are made to feel how
ignorant we really are.
[92] Pflüger's Archiv, vol. 44. p. 230-1.
[93] Naturally, as Schiff long ago pointed out (Lehrb.
D. Muskel-u. Nervenphysiologie, 1859, p. 213 ff.), the 'Rückenmarksseele,'
if it now exist, can have no higher sense-consciousness, for its incoming
currents are solely from the skin. But it may, in its dim way, both feel,
prefer, and desire. See, for the view favorable to the text: B.H. Lewes,
The Physiology of Common Life(1860), chap. IX. Goltz (Nervencentren des
Frosches, 1869, pp. 102-130) thinks that the frog's cord has no adaptive
power. This may be the case in such experiments as his, because the beheaded
frog's short span of life does not give it time to learn the new tricks
asked for. But Rosenthal (Biologisches Centralblatt, vol. IV. p. 247) and
Mendelssohn (Berlin Akad. Sitzungsberichte, 1885, p. 107) in their investigations
on the simple reflexes of the frog's cord, show that there is some adaptation
to new conditions, inasmuch as when usual paths of conduction are interrupted
by a cut, new paths are taken. According to Rosenthal, these grow more
pervious (i.e. require a smaller stimulus) in proportion as they are more
often traversed.
[94] Whether this evolution takes place through the
inheritance of habits acquired, or through the preservation of lucky variations,
is an alternative which we need not discuss here. We shall consider it
in the last chapter in the book. For our present purpose the modus operandi
of the evolution makes no difference, provided it be admitted to occur.
[95] See Schrader's Observations, loc. cit.